

A short polybasic segment between the two conserved domains of the β2a-subunit modulates the rate of inactivation of R-type calcium channel
- PMID: 22851179
- PMCID: PMC3463332
- DOI: 10.1074/jbc.M112.362509
A short polybasic segment between the two conserved domains of the β2a-subunit modulates the rate of inactivation of R-type calcium channel
Abstract
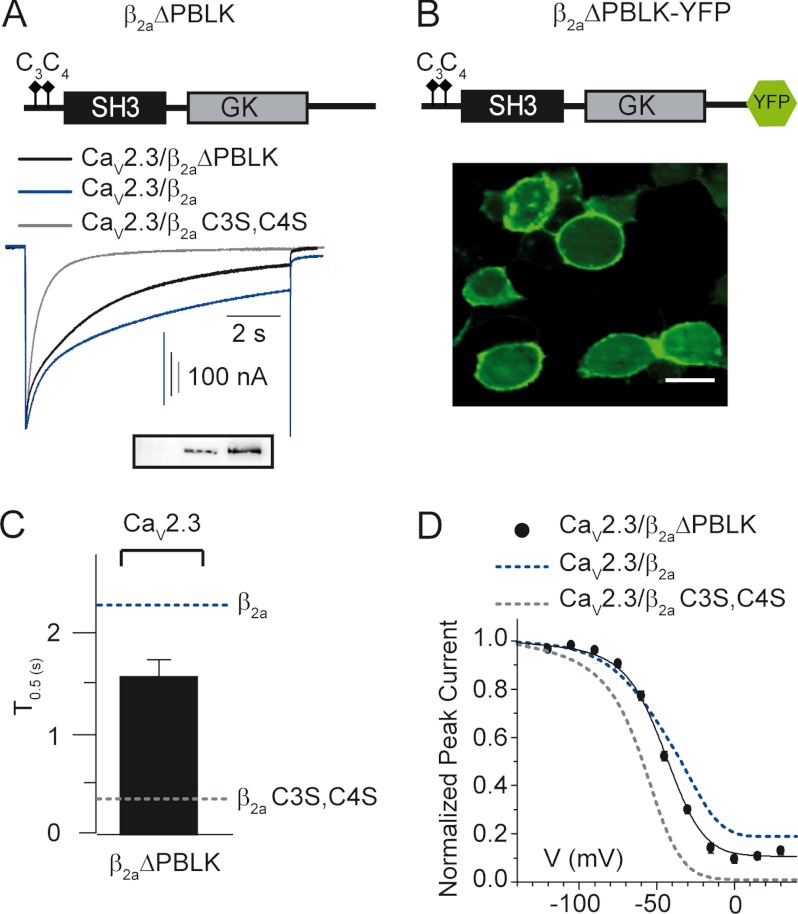
Besides opening and closing, high voltage-activated calcium channels transit to a nonconducting inactivated state from which they do not re-open unless the plasma membrane is repolarized. Inactivation is critical for temporal regulation of intracellular calcium signaling and prevention of a deleterious rise in calcium concentration. R-type high voltage-activated channels inactivate fully in a few hundred milliseconds when expressed alone. However, when co-expressed with a particular β-subunit isoform, β(2a), inactivation is partial and develops in several seconds. Palmitoylation of a unique di-cysteine motif at the N terminus anchors β(2a) to the plasma membrane. The current view is that membrane-anchored β(2a) immobilizes the channel inactivation machinery and confers slow inactivation phenotype. β-Subunits contain one Src homology 3 and one guanylate kinase domain, flanked by variable regions with unknown structures. Here, we identified a short polybasic segment at the boundary of the guanylate kinase domain that slows down channel inactivation without relocating a palmitoylation-deficient β(2a) to the plasma membrane. Substitution of the positively charged residues within this segment by alanine abolishes its slow inactivation-conferring phenotype. The linker upstream from the polybasic segment, but not the N- and C-terminal variable regions, masks the effect of this determinant. These results reveal a novel mechanism for inhibiting voltage-dependent inactivation of R-type calcium channels by the β(2a)-subunit that might involve electrostatic interactions with an unknown target on the channel's inactivation machinery or its modulatory components. They also suggest that intralinker interactions occlude the action of the polybasic segment and that its functional availability is regulated by the palmitoylated state of the β(2a)-subunit.
Figures






Similar articles
-
The HOOK-domain between the SH3 and the GK domains of Cavbeta subunits contains key determinants controlling calcium channel inactivation.Channels (Austin). 2007 Mar-Apr;1(2):92-101. doi: 10.4161/chan.4145. Epub 2007 Mar 15. Channels (Austin). 2007. PMID: 18690022
-
The guanylate kinase domain of the beta-subunit of voltage-gated calcium channels suffices to modulate gating.Proc Natl Acad Sci U S A. 2008 Sep 16;105(37):14198-203. doi: 10.1073/pnas.0806558105. Epub 2008 Sep 5. Proc Natl Acad Sci U S A. 2008. PMID: 18776052 Free PMC article.
-
The N terminus of a schistosome beta subunit regulates inactivation and current density of a Cav2 channel.J Biol Chem. 2010 Nov 12;285(46):35878-88. doi: 10.1074/jbc.M110.144725. Epub 2010 Sep 7. J Biol Chem. 2010. PMID: 20826800 Free PMC article.
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
-
Voltage-dependent calcium channels.Gen Physiol Biophys. 2005 Jun;24 Suppl 1:1-78. Gen Physiol Biophys. 2005. PMID: 16096350 Review.
Cited by
-
Characterization of the first honeybee Ca²⁺ channel subunit reveals two novel species- and splicing-specific modes of regulation of channel inactivation.Pflugers Arch. 2013 Jul;465(7):985-96. doi: 10.1007/s00424-013-1223-2. Epub 2013 Apr 16. Pflugers Arch. 2013. PMID: 23588376
-
Voltage-Gated Ca2+ Channels in Dopaminergic Substantia Nigra Neurons: Therapeutic Targets for Neuroprotection in Parkinson's Disease?Front Synaptic Neurosci. 2021 Feb 26;13:636103. doi: 10.3389/fnsyn.2021.636103. eCollection 2021. Front Synaptic Neurosci. 2021. PMID: 33716705 Free PMC article. Review.
-
Mapping the interaction surface between CaVβ and actin and its role in calcium channel clearance.Nat Commun. 2025 May 10;16(1):4352. doi: 10.1038/s41467-025-59548-x. Nat Commun. 2025. PMID: 40348749 Free PMC article.
-
Molecular basis of the PIP2-dependent regulation of CaV2.2 channel and its modulation by CaV β subunits.Elife. 2022 Nov 14;11:e69500. doi: 10.7554/eLife.69500. Elife. 2022. PMID: 36374183 Free PMC article.
-
The HOOK region of voltage-gated Ca2+ channel β subunits senses and transmits PIP2 signals to the gate.J Gen Physiol. 2017 Feb;149(2):261-276. doi: 10.1085/jgp.201611677. Epub 2017 Jan 13. J Gen Physiol. 2017. PMID: 28087621 Free PMC article.
References
-
- Clapham D. E. (2007) Calcium signaling. Cell 131, 1047–1058 - PubMed
-
- Catterall W. A. (2000) Structure and regulation of voltage-gated Ca2+ channels. Annu. Rev. Cell Dev. Biol. 16, 521–555 - PubMed
-
- Cens T., Rousset M., Leyris J. P., Fesquet P., Charnet P. (2006) Voltage- and calcium-dependent inactivation in high voltage-gated Ca2+ channels. Prog. Biophys. Mol. Biol. 90, 104–117 - PubMed
-
- Liang H., DeMaria C. D., Erickson M. G., Mori M. X., Alseikhan B. A., Yue D. T. (2003) Unified mechanisms of Ca2+ regulation across the Ca2+ channel family. Neuron 39, 951–960 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
