

Human H-ficolin inhibits replication of seasonal and pandemic influenza A viruses
- PMID: 22851708
- PMCID: PMC3650843
- DOI: 10.4049/jimmunol.1103786
Human H-ficolin inhibits replication of seasonal and pandemic influenza A viruses
Abstract
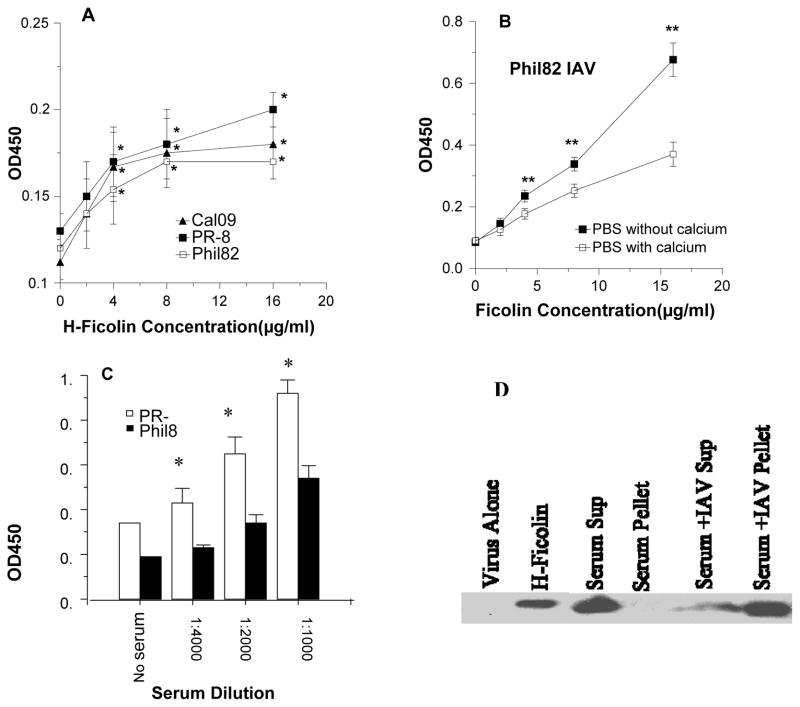
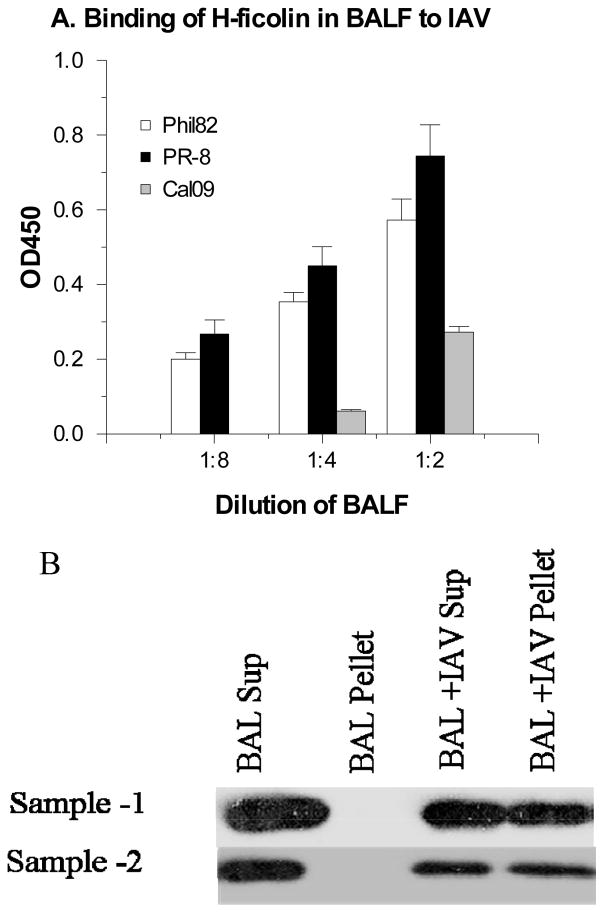
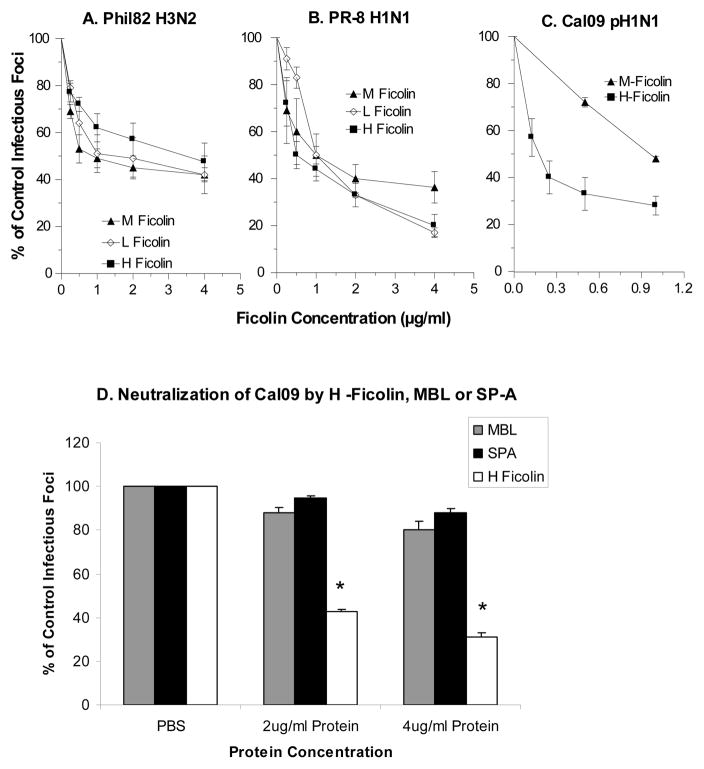
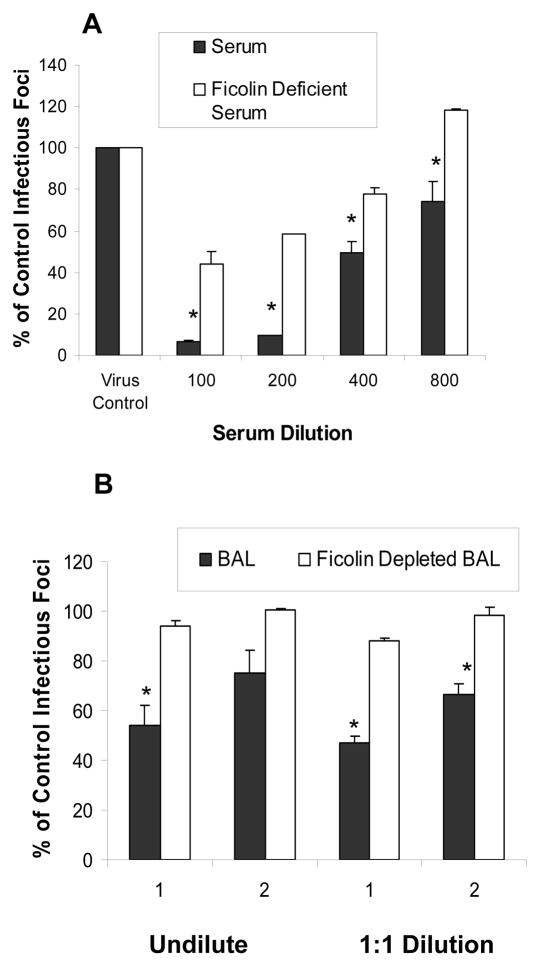
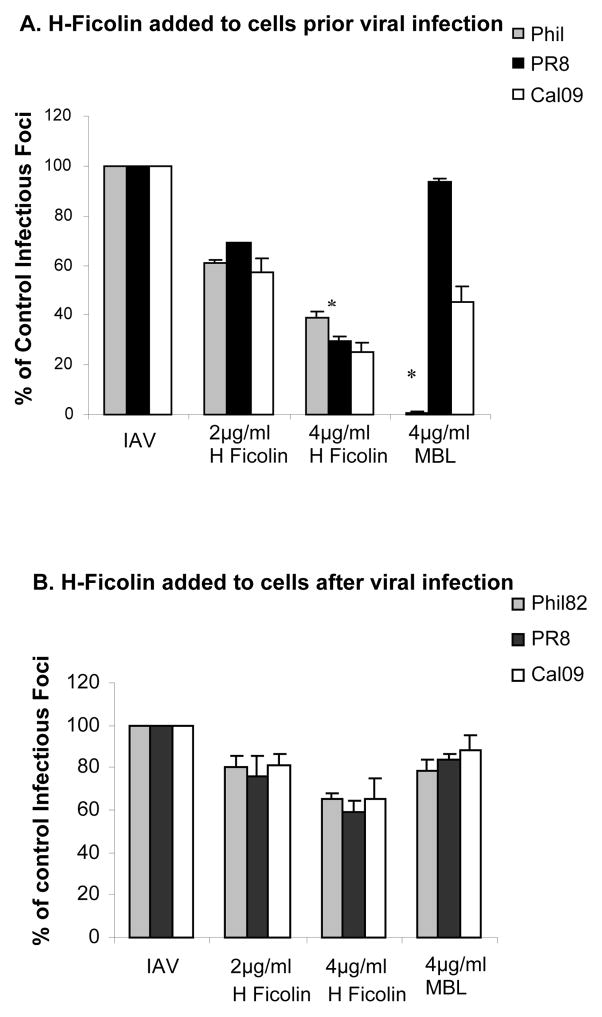
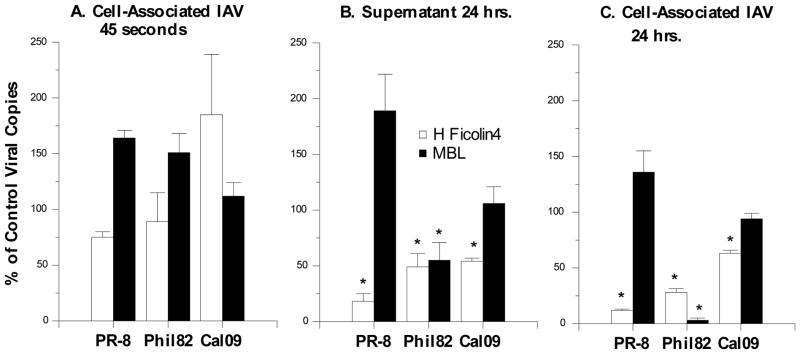
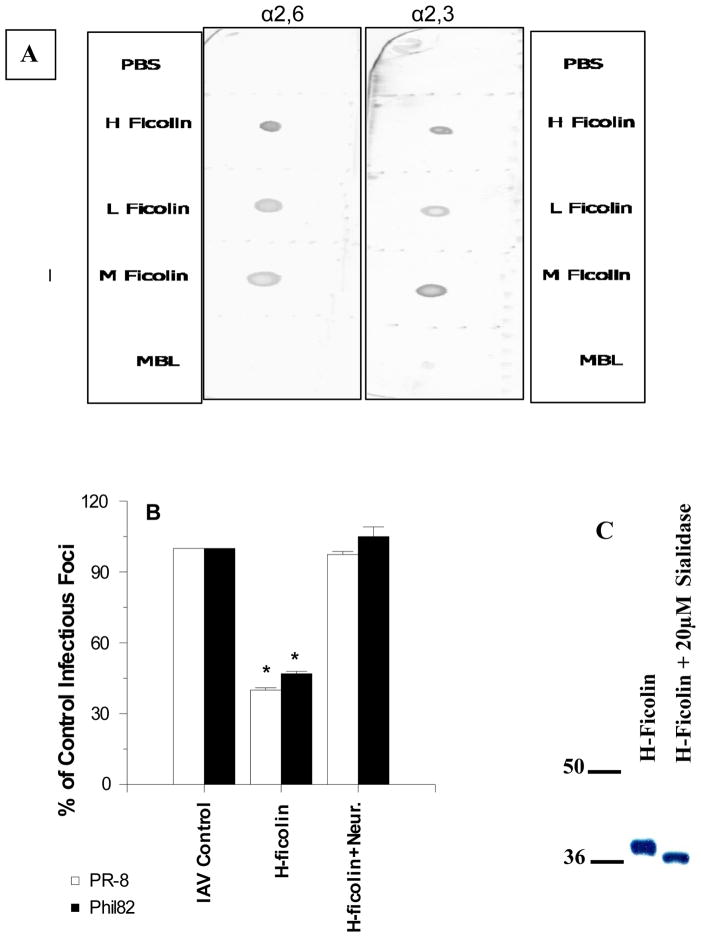
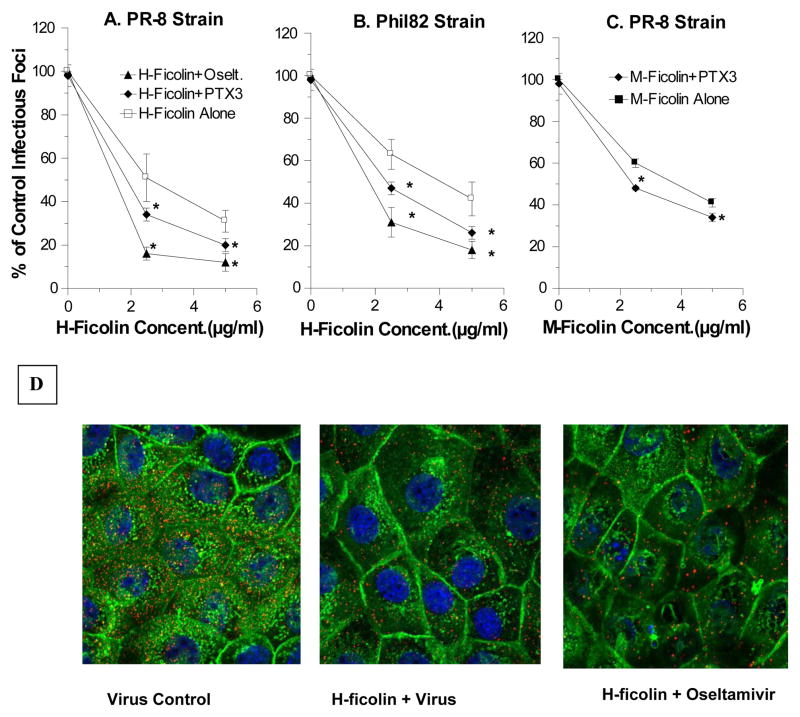
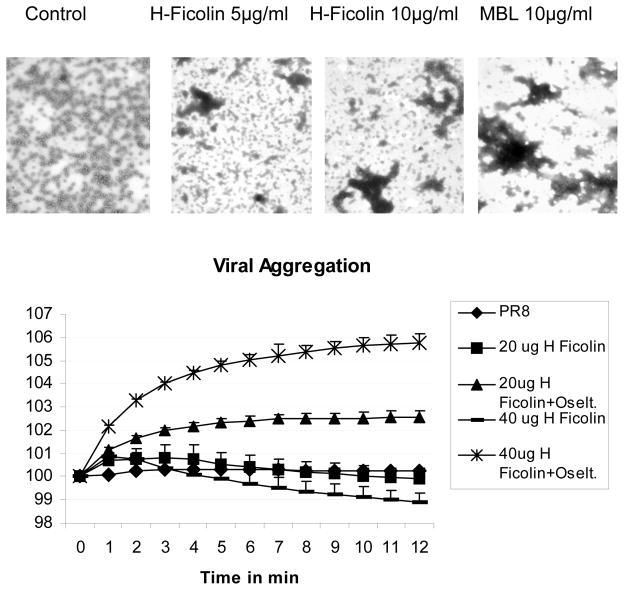
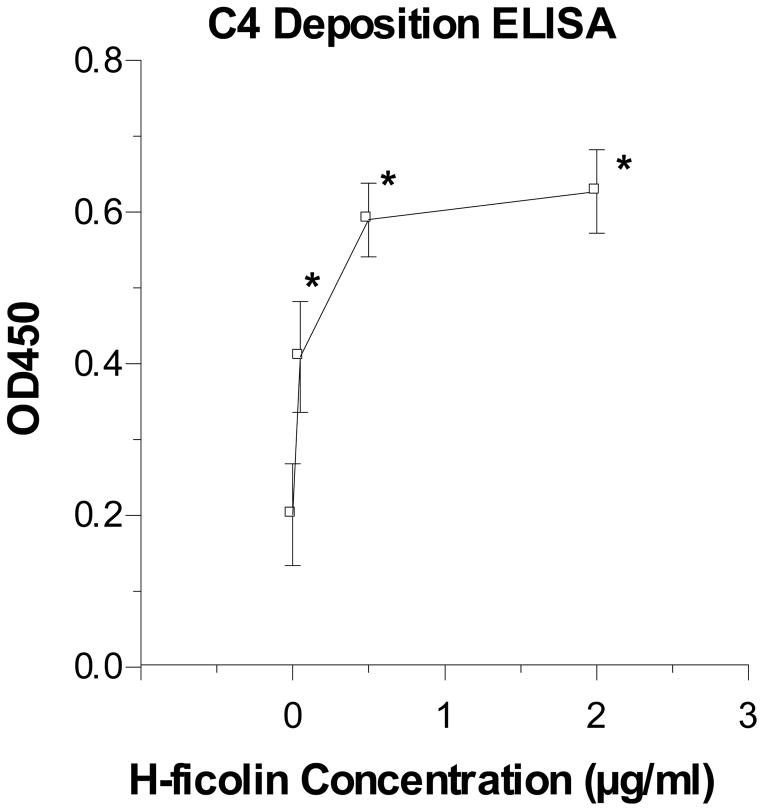
The collectins have been shown to have a role in host defense against influenza A virus (IAV) and other significant viral pathogens (e.g., HIV). The ficolins are a related group of innate immune proteins that are present at relatively high concentrations in serum, but also in respiratory secretions; however, there has been little study of the role of ficolins in viral infection. In this study, we demonstrate that purified recombinant human H-ficolin and H-ficolin in human serum and bronchoalveolar lavage fluid bind to IAV and inhibit viral infectivity and hemagglutination activity in vitro. Removal of ficolins from human serum or bronchoalveolar lavage fluid reduces their antiviral activity. Inhibition of IAV did not involve the calcium-dependent lectin activity of H-ficolin. We demonstrate that H-ficolin is sialylated and that removal of sialic acid abrogates IAV inhibition, while addition of the neuraminidase inhibitor oseltamivir potentiates neutralization, hemagglutinin inhibition, and viral aggregation caused by H-ficolin. Pandemic and mouse-adapted strains of IAV are generally not inhibited by the collectins surfactant protein D or mannose binding lectin because of a paucity of glycan attachments on the hemagglutinin of these strains. In contrast, H-ficolin inhibited both the mouse-adapted PR-8 H1N1 strain and a pandemic H1N1 strain from 2009. H-ficolin also fixed complement to a surface coated with IAV. These findings suggest that H-ficolin contributes to host defense against IAV.
Figures
Similar articles
-
Collectins, H-ficolin and LL-37 reduce influence viral replication in human monocytes and modulate virus-induced cytokine production.Innate Immun. 2017 Jan;23(1):77-88. doi: 10.1177/1753425916678470. Epub 2016 Nov 17. Innate Immun. 2017. PMID: 27856789
-
Innate defense against influenza A virus: activity of human neutrophil defensins and interactions of defensins with surfactant protein D.J Immunol. 2006 Jun 1;176(11):6962-72. doi: 10.4049/jimmunol.176.11.6962. J Immunol. 2006. PMID: 16709857
-
Full-length human surfactant protein A inhibits influenza A virus infection of A549 lung epithelial cells: A recombinant form containing neck and lectin domains promotes infectivity.Immunobiology. 2019 May;224(3):408-418. doi: 10.1016/j.imbio.2019.02.006. Epub 2019 Feb 11. Immunobiology. 2019. PMID: 30954271
-
H3N2 influenza viruses in humans: Viral mechanisms, evolution, and evaluation.Hum Vaccin Immunother. 2018;14(8):1840-1847. doi: 10.1080/21645515.2018.1462639. Epub 2018 May 14. Hum Vaccin Immunother. 2018. PMID: 29641358 Free PMC article. Review.
-
The Role and Molecular Mechanism of Action of Surfactant Protein D in Innate Host Defense Against Influenza A Virus.Front Immunol. 2018 Jun 13;9:1368. doi: 10.3389/fimmu.2018.01368. eCollection 2018. Front Immunol. 2018. PMID: 29951070 Free PMC article. Review.
Cited by
-
Human lectins and their roles in viral infections.Molecules. 2015 Jan 29;20(2):2229-71. doi: 10.3390/molecules20022229. Molecules. 2015. PMID: 25642836 Free PMC article. Review.
-
Alzheimer's associated β-amyloid protein inhibits influenza A virus and modulates viral interactions with phagocytes.PLoS One. 2014 Jul 2;9(7):e101364. doi: 10.1371/journal.pone.0101364. eCollection 2014. PLoS One. 2014. PMID: 24988208 Free PMC article.
-
Neuraminidase-mediated, NKp46-dependent immune-evasion mechanism of influenza viruses.Cell Rep. 2013 Apr 25;3(4):1044-50. doi: 10.1016/j.celrep.2013.03.034. Epub 2013 Apr 18. Cell Rep. 2013. PMID: 23602571 Free PMC article.
-
Biological activities of 'noninfectious' influenza A virus particles.Future Virol. 2014 Jan;9(1):41-51. doi: 10.2217/fvl.13.118. Future Virol. 2014. PMID: 25067941 Free PMC article.
-
Influenza virus N-linked glycosylation and innate immunity.Biosci Rep. 2019 Jan 8;39(1):BSR20171505. doi: 10.1042/BSR20171505. Print 2019 Jan 31. Biosci Rep. 2019. PMID: 30552137 Free PMC article. Review.
References
-
- Thiel S. Complement activating soluble pattern recognition molecules with collagen-like regions, mannan-binding lectin, ficolins and associated proteins. Mol Immunol. 2007;44:3875–3888. - PubMed
-
- Garred P, Borregaard N. The ficolins. J Innate Immun. 2009;2:1–2. - PubMed
-
- Akaiwa M, Yae Y, Sugimoto R, Suzuki SO, Iwaki T, Izuhara K, Hamasaki N. Hakata antigen, a new member of the ficolin/opsonin p35 family, is a novel human lectin secreted into bronchus/alveolus and bile. J Histochem Cytochem. 1999;47:777–786. - PubMed
-
- Sallenbach S, Thiel S, Aebi C, Otth M, Bigler S, Jensenius JC, Schlapbach LJ, Ammann RA. Serum concentrations of lectin-pathway components in healthy neonates, children and adults: mannan-binding lectin (MBL), M-, L-, and H-ficolin, and MBL-associated serine protease-2 (MASP-2) Pediatr Allergy Immunol. 2011;22:424–430. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
