

Bioinformatic cis-element analyses performed in Arabidopsis and rice disclose bZIP- and MYB-related binding sites as potential AuxRE-coupling elements in auxin-mediated transcription
- PMID: 22852874
- PMCID: PMC3438128
- DOI: 10.1186/1471-2229-12-125
Bioinformatic cis-element analyses performed in Arabidopsis and rice disclose bZIP- and MYB-related binding sites as potential AuxRE-coupling elements in auxin-mediated transcription
Abstract
Background: In higher plants, a diverse array of developmental and growth-related processes is regulated by the plant hormone auxin. Recent publications have proposed that besides the well-characterized Auxin Response Factors (ARFs) that bind Auxin Response Elements (AuxREs), also members of the bZIP- and MYB-transcription factor (TF) families participate in transcriptional control of auxin-regulated genes via bZIP Response Elements (ZREs) or Myb Response Elements (MREs), respectively.
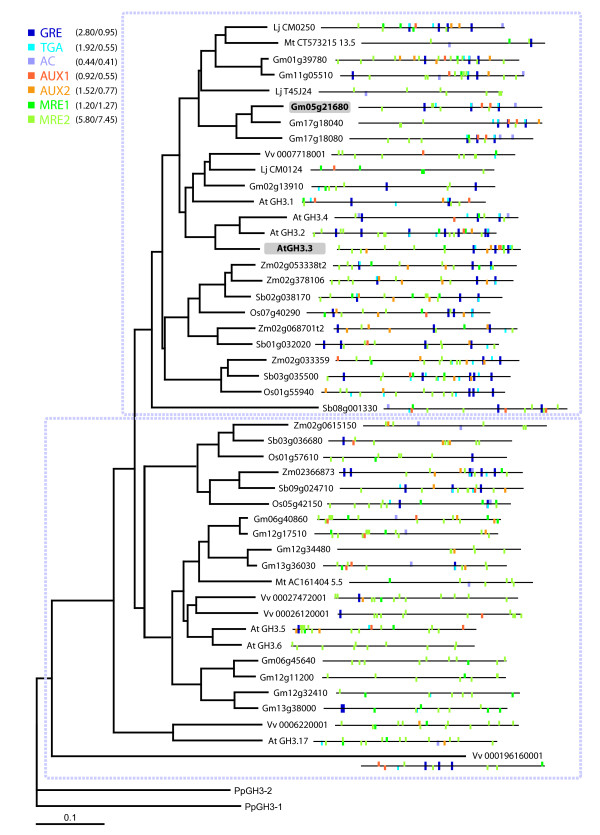
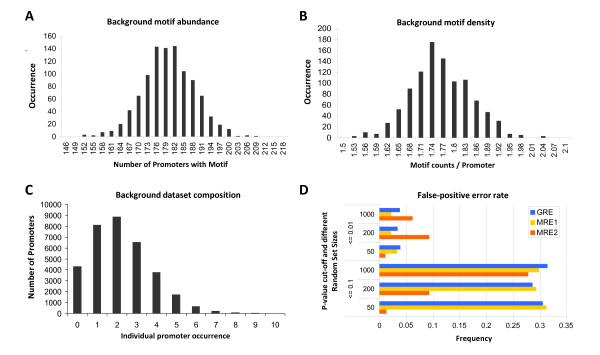
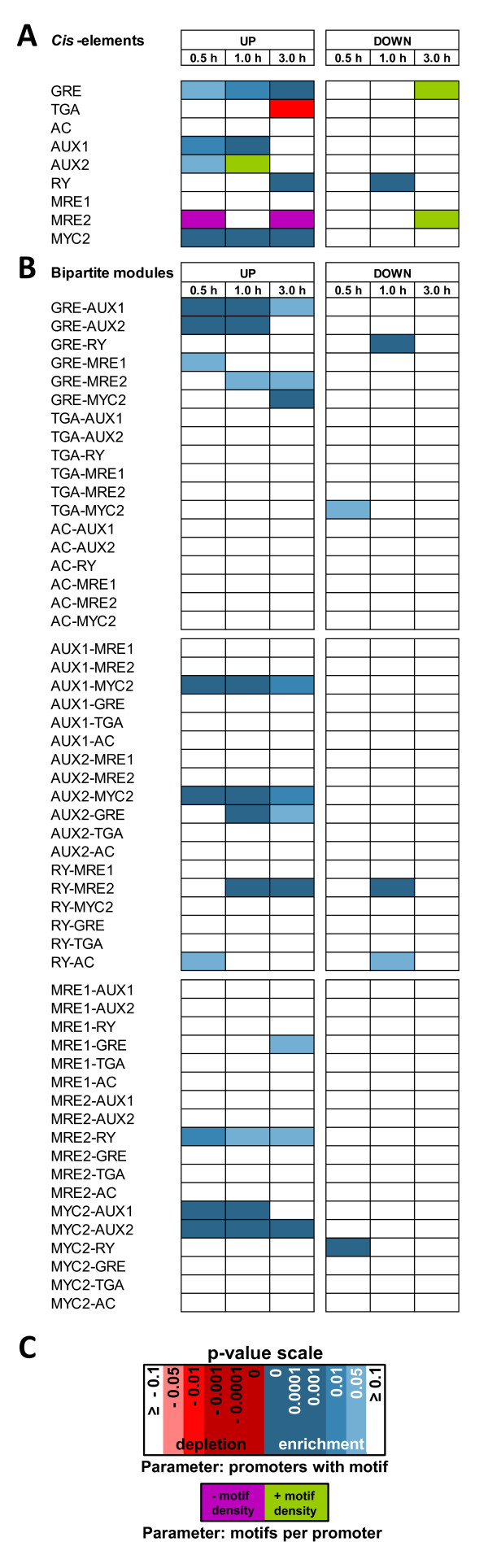
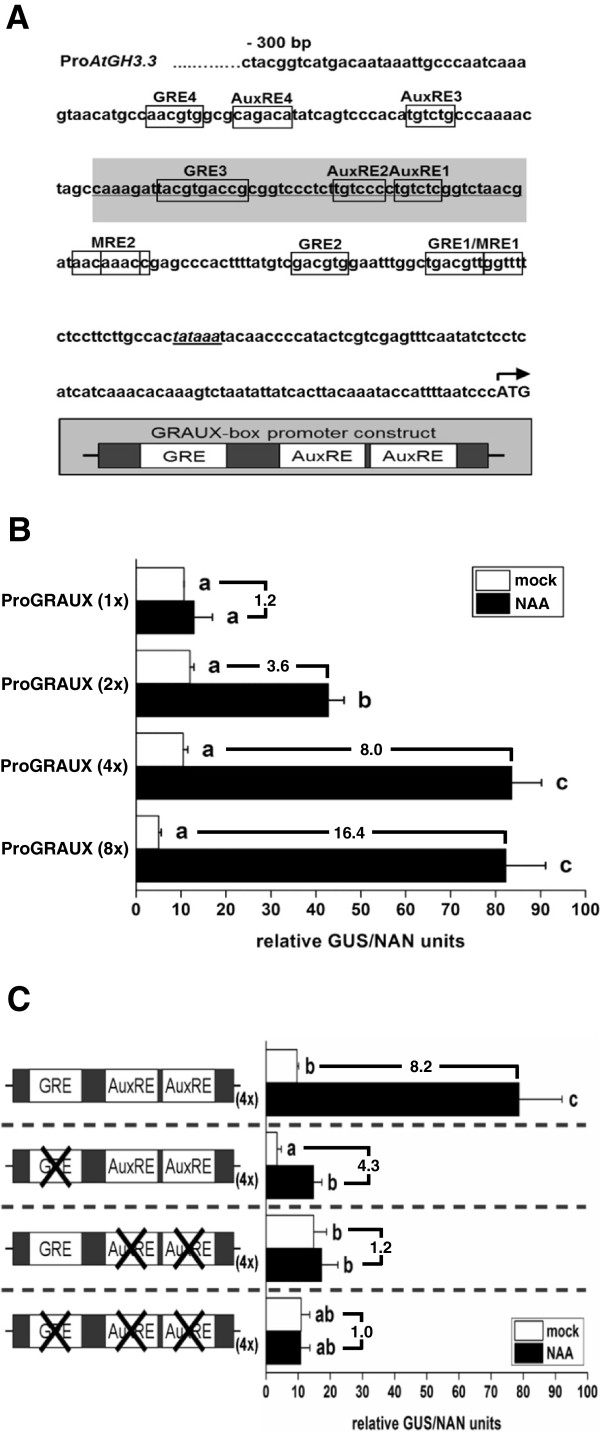
Results: Applying a novel bioinformatic algorithm, we demonstrate on a genome-wide scale that singular motifs or composite modules of AuxREs, ZREs, MREs but also of MYC2 related elements are significantly enriched in promoters of auxin-inducible genes. Despite considerable, species-specific differences in the genome structure in terms of the GC content, this enrichment is generally conserved in dicot (Arabidopsis thaliana) and monocot (Oryza sativa) model plants. Moreover, an enrichment of defined composite modules has been observed in selected auxin-related gene families. Consistently, a bipartite module, which encompasses a bZIP-associated G-box Related Element (GRE) and an AuxRE motif, has been found to be highly enriched. Making use of transient reporter studies in protoplasts, these findings were experimentally confirmed, demonstrating that GREs functionally interact with AuxREs in regulating auxin-mediated transcription.
Conclusions: Using genome-wide bioinformatic analyses, evolutionary conserved motifs have been defined which potentially function as AuxRE-dependent coupling elements to establish auxin-specific expression patterns. Based on these findings, experimental approaches can be designed to broaden our understanding of combinatorial, auxin-controlled gene regulation.
Figures

References
-
- Bennett T, Scheres B. Root development-two meristems for the price of one? Curr Top Dev Biol. 2010;91:67–102. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
