

A microRNA network regulates expression and biosynthesis of wild-type and DeltaF508 mutant cystic fibrosis transmembrane conductance regulator
- PMID: 22853952
- PMCID: PMC3421220
- DOI: 10.1073/pnas.1210906109
A microRNA network regulates expression and biosynthesis of wild-type and DeltaF508 mutant cystic fibrosis transmembrane conductance regulator
Abstract
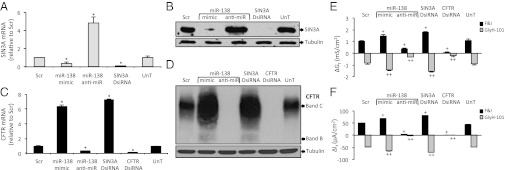
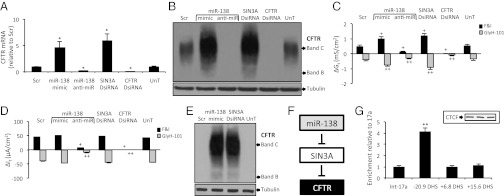
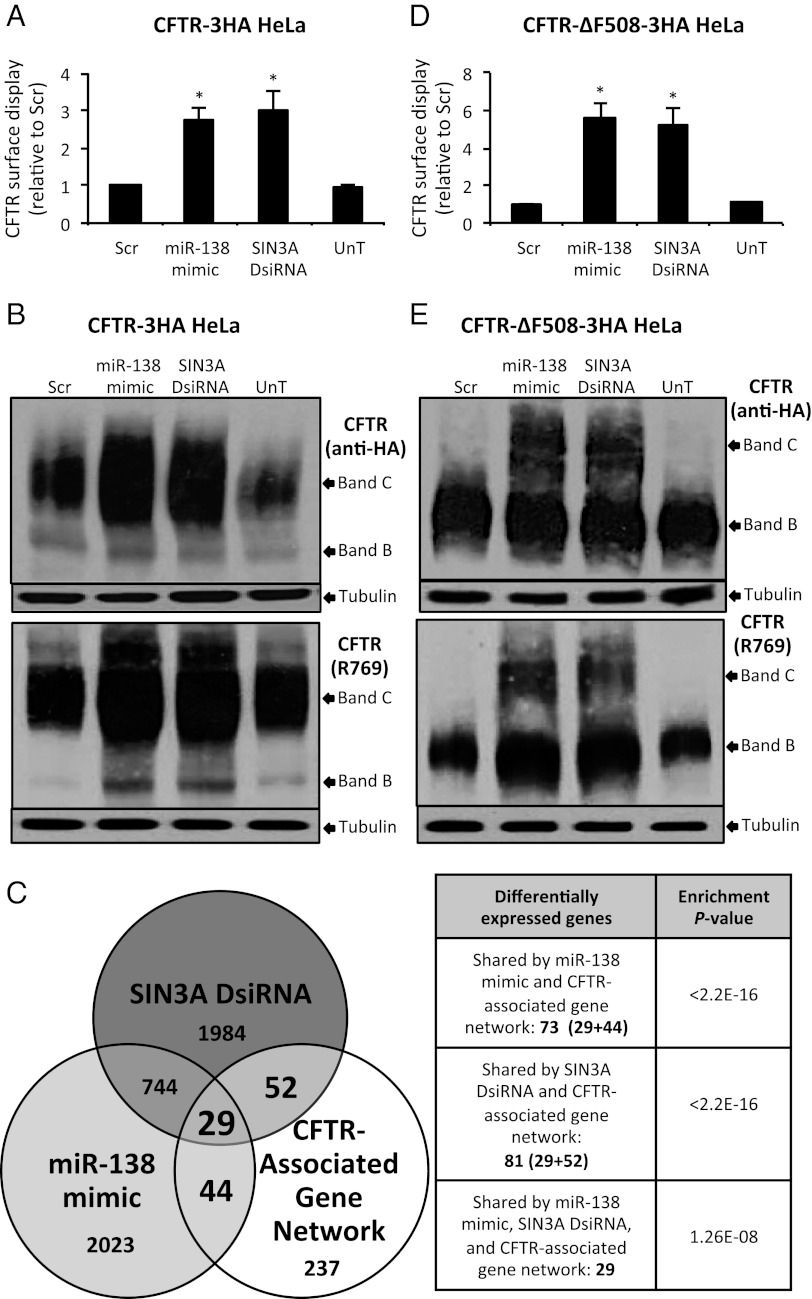
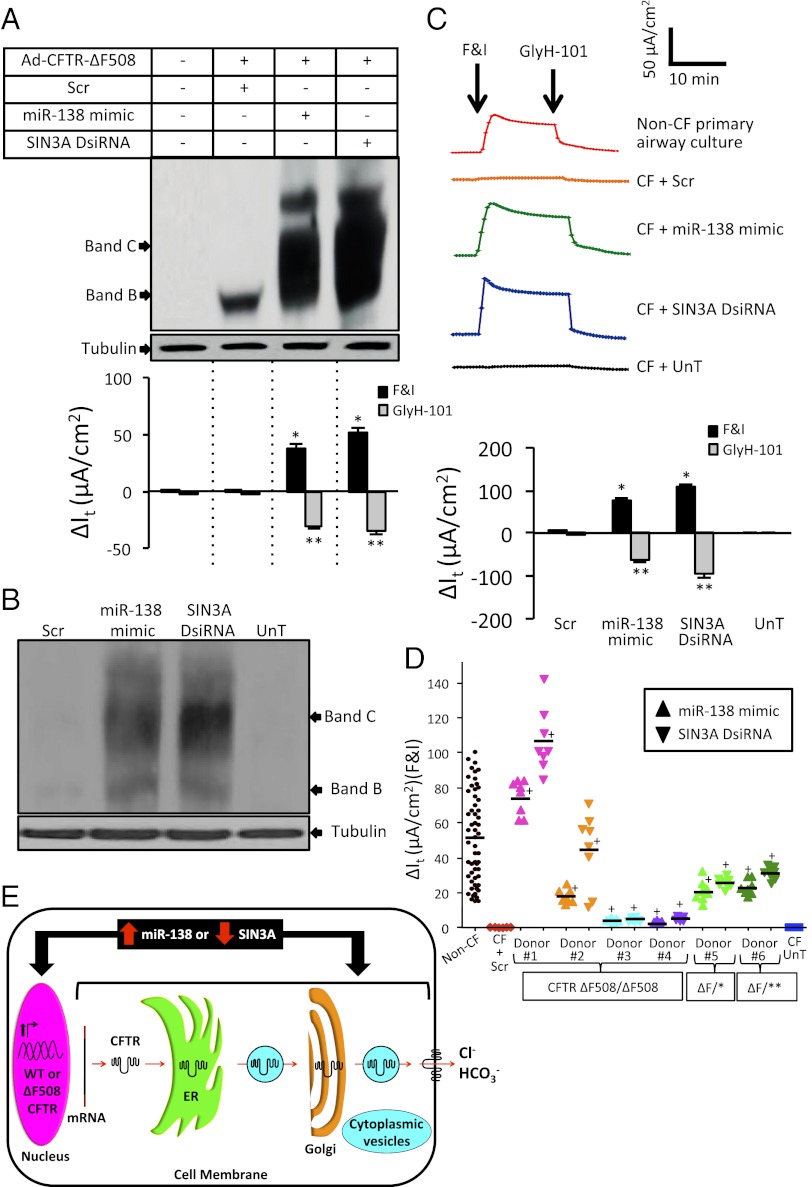
Production of functional proteins requires multiple steps, including gene transcription and posttranslational processing. MicroRNAs (miRNAs) can regulate individual stages of these processes. Despite the importance of the cystic fibrosis transmembrane conductance regulator (CFTR) channel for epithelial anion transport, how its expression is regulated remains uncertain. We discovered that miRNA-138 regulates CFTR expression through its interactions with the transcriptional regulatory protein SIN3A. Treating airway epithelia with an miR-138 mimic increased CFTR mRNA and also enhanced CFTR abundance and transepithelial Cl(-) permeability independent of elevated mRNA levels. An miR-138 anti-miR had the opposite effects. Importantly, miR-138 altered the expression of many genes encoding proteins that associate with CFTR and may influence its biosynthesis. The most common CFTR mutation, ΔF508, causes protein misfolding, protein degradation, and cystic fibrosis. Remarkably, manipulating the miR-138 regulatory network also improved biosynthesis of CFTR-ΔF508 and restored Cl(-) transport to cystic fibrosis airway epithelia. This miRNA-regulated network directs gene expression from the chromosome to the cell membrane, indicating that an individual miRNA can control a cellular process more broadly than recognized previously. This discovery also provides therapeutic avenues for restoring CFTR function to cells affected by the most common cystic fibrosis mutation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
