

The antisense RNA As1_flv4 in the Cyanobacterium Synechocystis sp. PCC 6803 prevents premature expression of the flv4-2 operon upon shift in inorganic carbon supply
- PMID: 22854963
- PMCID: PMC3460422
- DOI: 10.1074/jbc.M112.391755
The antisense RNA As1_flv4 in the Cyanobacterium Synechocystis sp. PCC 6803 prevents premature expression of the flv4-2 operon upon shift in inorganic carbon supply
Abstract
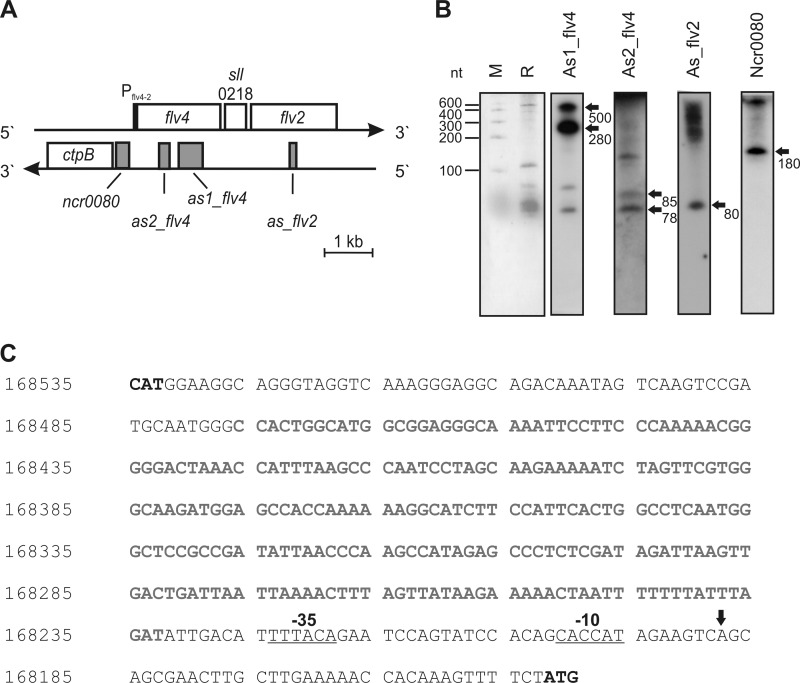
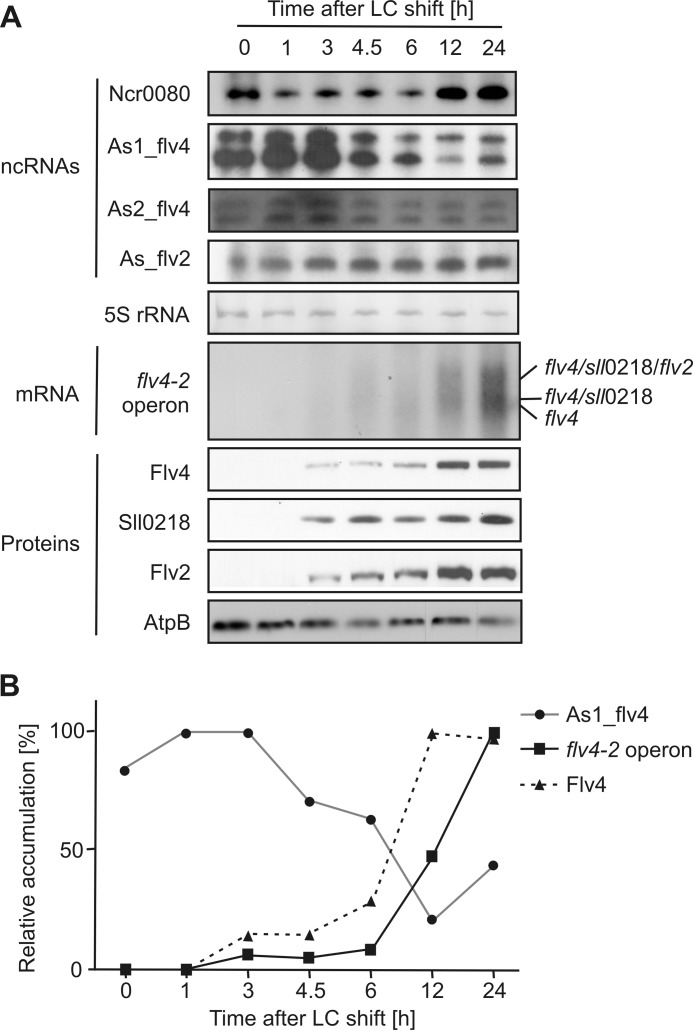
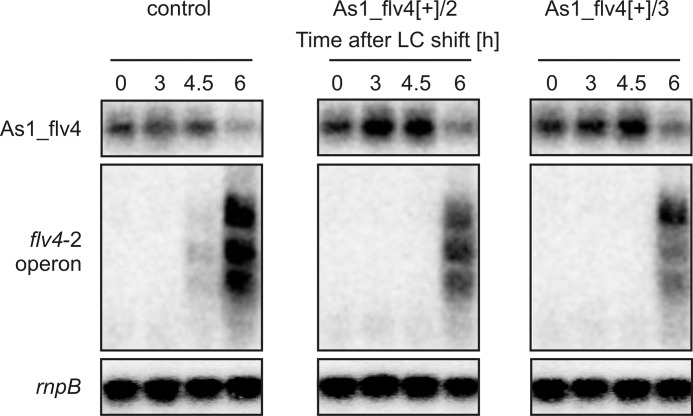
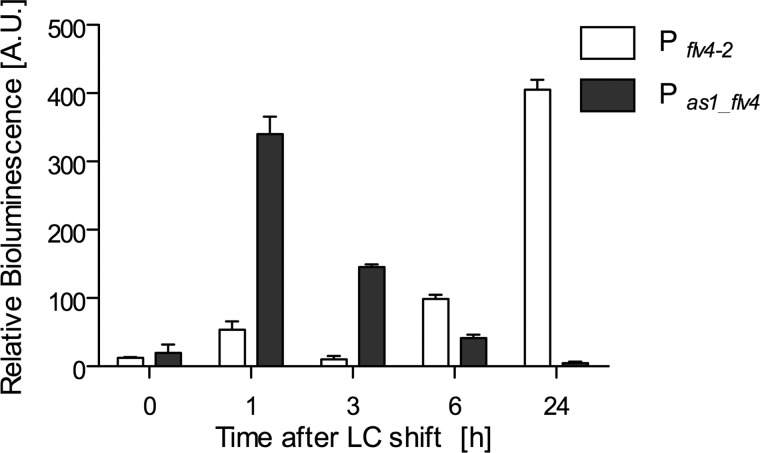
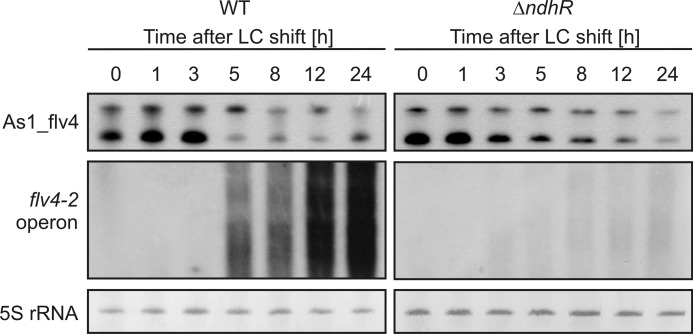
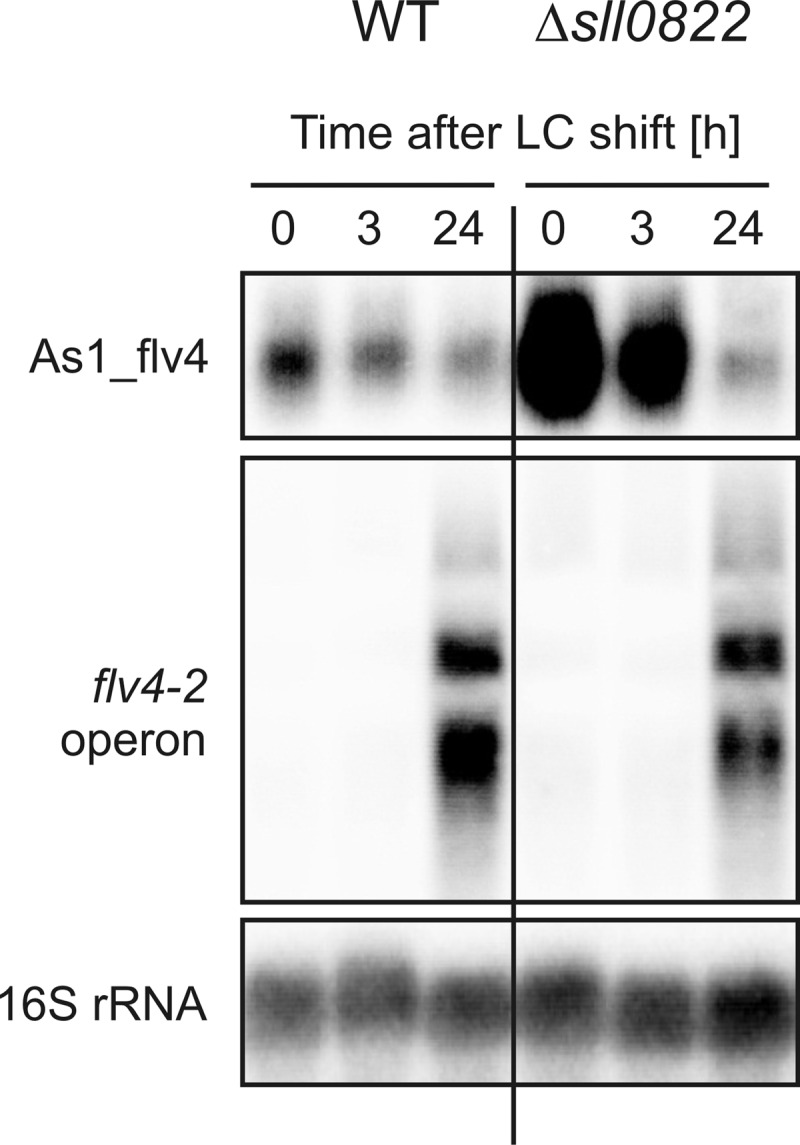
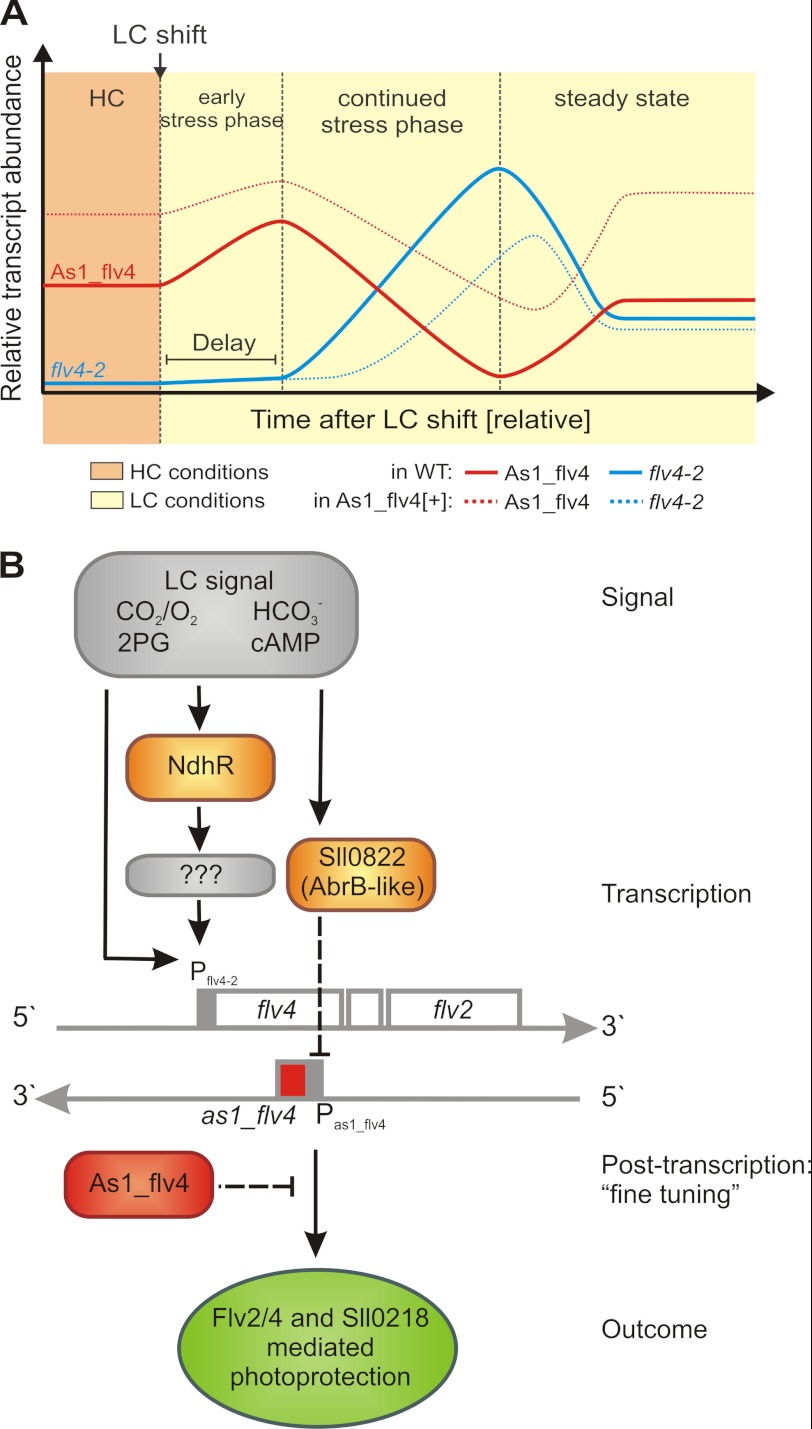
The functional relevance of natural cis-antisense transcripts is mostly unknown. Here we have characterized the association of three antisense RNAs and one intergenically encoded noncoding RNA with an operon that plays a crucial role in photoprotection of photosystem II under low carbon conditions in the cyanobacterium Synechocystis sp. PCC 6803. Cyanobacteria show strong gene expression dynamics in response to a shift of cells from high carbon to low levels of inorganic carbon (C(i)), but the regulatory mechanisms are poorly understood. Among the most up-regulated genes in Synechocystis are flv4, sll0218, and flv2, which are organized in the flv4-2 operon. The flavodiiron proteins encoded by this operon open up an alternative electron transfer route, likely starting from the Q(B) site in photosystem II, under photooxidative stress conditions. Our expression analysis of cells shifted from high carbon to low carbon demonstrated an inversely correlated transcript accumulation of the flv4-2 operon mRNA and one antisense RNA to flv4, designated as As1_flv4. Overexpression of As1_flv4 led to a decrease in flv4-2 mRNA. The promoter activity of as1_flv4 was transiently stimulated by C(i) limitation and negatively regulated by the AbrB-like transcription regulator Sll0822, whereas the flv4-2 operon was positively regulated by the transcription factor NdhR. The results indicate that the tightly regulated antisense RNA As1_flv4 establishes a transient threshold for flv4-2 expression in the early phase after a change in C(i) conditions. Thus, it prevents unfavorable synthesis of the proteins from the flv4-2 operon.
Figures
References
-
- De Lucia F., Dean C. (2011) Long non-coding RNAs and chromatin regulation. Curr. Opin. Plant Biol. 14, 168–173 - PubMed
-
- Zhang X., Xia J., Lii Y. E., Barrera-Figueroa B. E., Zhou X., Gao S., Lu L., Niu D., Chen Z., Leung C., Wong T., Zhang H., Guo J., Li Y., Liu R., Liang W., Zhu J. K., Zhang W., Jin H. (2012) Genome-wide analysis of plant nat-siRNAs reveals insights into their distribution, biogenesis and function. Genome Biol. 13, R20. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
