

Cell type-specific genomics of Drosophila neurons
- PMID: 22855560
- PMCID: PMC3479168
- DOI: 10.1093/nar/gks671
Cell type-specific genomics of Drosophila neurons
Abstract
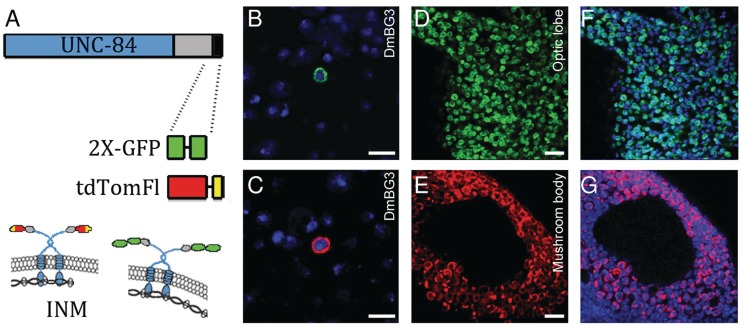
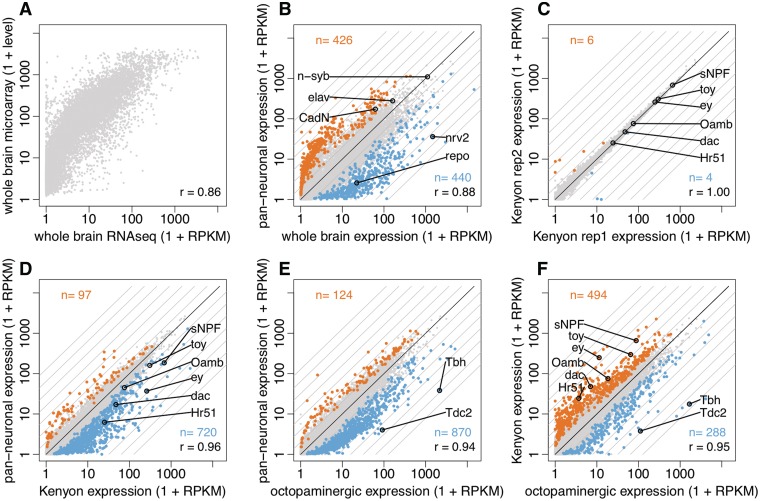
Many tools are available to analyse genomes but are often challenging to use in a cell type-specific context. We have developed a method similar to the isolation of nuclei tagged in a specific cell type (INTACT) technique [Deal,R.B. and Henikoff,S. (2010) A simple method for gene expression and chromatin profiling of individual cell types within a tissue. Dev. Cell, 18, 1030-1040; Steiner,F.A., Talbert,P.B., Kasinathan,S., Deal,R.B. and Henikoff,S. (2012) Cell-type-specific nuclei purification from whole animals for genome-wide expression and chromatin profiling. Genome Res., doi:10.1101/gr.131748.111], first developed in plants, for use in Drosophila neurons. We profile gene expression and histone modifications in Kenyon cells and octopaminergic neurons in the adult brain. In addition to recovering known gene expression differences, we also observe significant cell type-specific chromatin modifications. In particular, a small subset of differentially expressed genes exhibits a striking anti-correlation between repressive and activating histone modifications. These genes are enriched for transcription factors, recovering those known to regulate mushroom body identity and predicting analogous regulators of octopaminergic neurons. Our results suggest that applying INTACT to specific neuronal populations can illuminate the transcriptional regulatory networks that underlie neuronal cell identity.
Figures




Similar articles
-
Genome-wide analysis of the binding of the Hox protein Ultrabithorax and the Hox cofactor Homothorax in Drosophila.PLoS One. 2011 Apr 5;6(4):e14778. doi: 10.1371/journal.pone.0014778. PLoS One. 2011. PMID: 21483667 Free PMC article.
-
Functional genomics of histone modification and non-histone chromosomal proteins using the polytene chromosomes of Drosophila.Methods. 2006 Dec;40(4):360-4. doi: 10.1016/j.ymeth.2006.09.001. Methods. 2006. PMID: 17101449
-
Cell-type-specific nuclei purification from whole animals for genome-wide expression and chromatin profiling.Genome Res. 2012 Apr;22(4):766-77. doi: 10.1101/gr.131748.111. Epub 2012 Jan 4. Genome Res. 2012. PMID: 22219512 Free PMC article.
-
DamID: a methylation-based chromatin profiling approach.Methods Mol Biol. 2009;567:155-69. doi: 10.1007/978-1-60327-414-2_11. Methods Mol Biol. 2009. PMID: 19588092 Review.
-
Interplay between different epigenetic modifications and mechanisms.Adv Genet. 2010;70:101-41. doi: 10.1016/B978-0-12-380866-0.60005-8. Adv Genet. 2010. PMID: 20920747 Review.
Cited by
-
Cell-type-specific profiling of gene expression and chromatin binding without cell isolation: assaying RNA Pol II occupancy in neural stem cells.Dev Cell. 2013 Jul 15;26(1):101-12. doi: 10.1016/j.devcel.2013.05.020. Epub 2013 Jun 20. Dev Cell. 2013. PMID: 23792147 Free PMC article.
-
Isolation and RNA sequencing of single nuclei from Drosophila tissues.STAR Protoc. 2022 May 20;3(2):101417. doi: 10.1016/j.xpro.2022.101417. eCollection 2022 Jun 17. STAR Protoc. 2022. PMID: 35620068 Free PMC article.
-
Insights from intoxicated Drosophila.Alcohol. 2019 Feb;74:21-27. doi: 10.1016/j.alcohol.2018.03.004. Epub 2018 Mar 21. Alcohol. 2019. PMID: 29980341 Free PMC article. Review.
-
Identification of sensory hair-cell transcripts by thiouracil-tagging in zebrafish.BMC Genomics. 2015 Oct 23;16:842. doi: 10.1186/s12864-015-2072-5. BMC Genomics. 2015. PMID: 26494580 Free PMC article.
-
Temporally and spatially restricted gene expression profiling.Curr Genomics. 2014 Aug;15(4):278-92. doi: 10.2174/1389202915666140602230106. Curr Genomics. 2014. PMID: 25132798 Free PMC article.
References
-
- Davidson EH. The regulatory genome: gene regulatory networks in development and evolution. Amsterdam: Elsevier, Acad. Press; 2006.
-
- Hobert O. Regulation of terminal differentiation programs in the nervous system. Annual Ann. Review of Cell and Developmental. Biology Biol. 2011;27:681–696. - PubMed
-
- Emmert-Buck MR, Bonner RF, Smith PD, et al. Laser capture microdissection. Science. 1996;274:998–1001. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
