

Strain background influences neurotoxicity and behavioral abnormalities in mice expressing the tetracycline transactivator
- PMID: 22855807
- PMCID: PMC3431916
- DOI: 10.1523/JNEUROSCI.0893-12.2012
Strain background influences neurotoxicity and behavioral abnormalities in mice expressing the tetracycline transactivator
Abstract
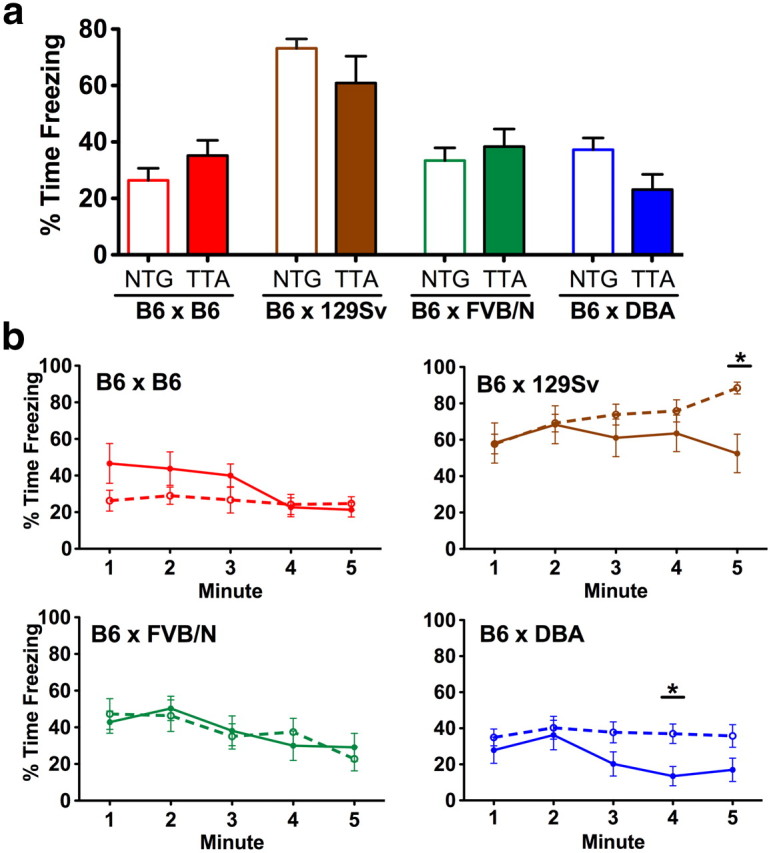
The tet-off system has been widely used to create transgenic models of neurological disorders including Alzheimer's, Parkinson's, Huntington's, and prion disease. The utility of this system lies in the assumption that the tetracycline transactivator (TTA) acts as an inert control element and does not contribute to phenotypes under study. Here we report that neuronal expression of TTA can affect hippocampal cytoarchitecture and behavior in a strain-dependent manner. While studying neurodegeneration in two tet-off Alzheimer's disease models, we unexpectedly discovered neuronal loss within the dentate gyrus of single transgenic TTA controls. Granule neurons appeared most sensitive to TTA exposure during postnatal development, and doxycycline treatment during this period was neuroprotective. TTA-induced degeneration could be rescued by moving the transgene onto a congenic C57BL/6J background and recurred on reintroduction of either CBA or C3H/He backgrounds. Quantitative trait analysis of B6C3 F2 TTA mice identified a region on Chromosome 14 that contains a major modifier of the neurodegenerative phenotype. Although B6 mice were resistant to degeneration, they were not ideal for cognitive testing. F1 offspring of TTA C57BL/6J and 129X1/SvJ, FVB/NJ, or DBA/1J showed improved spatial learning, but TTA expression caused subtle differences in contextual fear conditioning on two of these backgrounds, indicating that strain and genotype can interact independently under different behavioral settings. All model systems have limitations that should be recognized and mitigated where possible; our findings stress the importance of mapping the effects caused by TTA alone when working with tet-off models.
Conflict of interest statement
The authors declare no financial conflicts of interest.
Figures









References
-
- Altman J, Bayer SA. Mosaic organization of the hippocampal neuroepithelium and the multiple germinal sources of dentate granule cells. J Comp Neurol. 1990a;301:325–342. - PubMed
-
- Altman J, Bayer SA. Migration and distribution of two populations of hippocampal granule cell precursors during the perinatal and postnatal periods. J Comp Neurol. 1990b;301:365–381. - PubMed
-
- Alvarez-Saavedra M, Sáez MA, Kang D, Zoghbi HY, Young JI. Cell-specific expression of wild-type MeCP2 in mouse models of Rett syndrome yields insight about pathogenesis. Hum Mol Genet. 2007;16:2315–2325. - PubMed
-
- Barton MD, Dunlop JW, Psaltis G, Kulik J, DeGennaro L, Kwak SP. Modified GFAP promoter auto-regulates tet-activator expression for increased transactivation and reduced tTA-associated toxicity. Brain Res Mol Brain Res. 2002;101:71–81. - PubMed
-
- Bayer SA, Yackel JW, Puri PS. Neurons in the rat dentate gyrus granular layer substantially increase during juvenile and adult life. Science. 1982;216:890–892. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous