

Molecular basis for specific regulation of neuronal kinesin-3 motors by doublecortin family proteins
- PMID: 22857951
- PMCID: PMC3549492
- DOI: 10.1016/j.molcel.2012.06.025
Molecular basis for specific regulation of neuronal kinesin-3 motors by doublecortin family proteins
Abstract
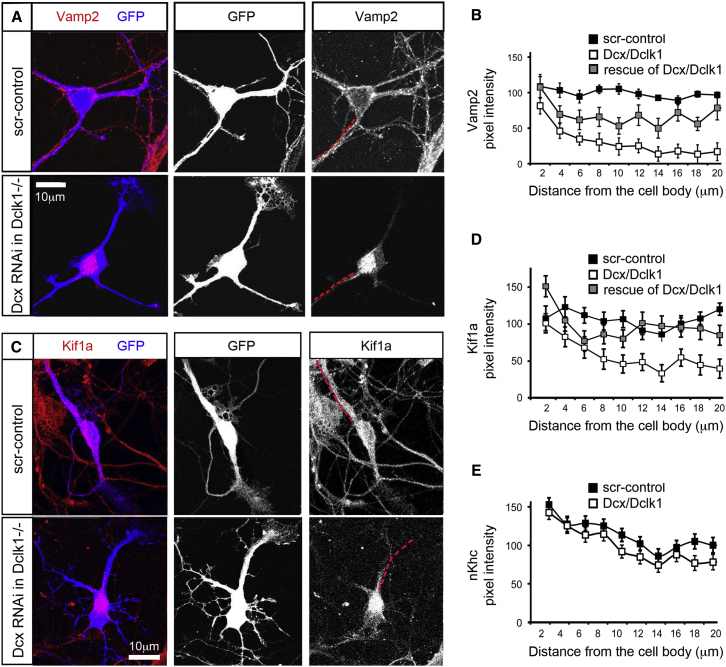
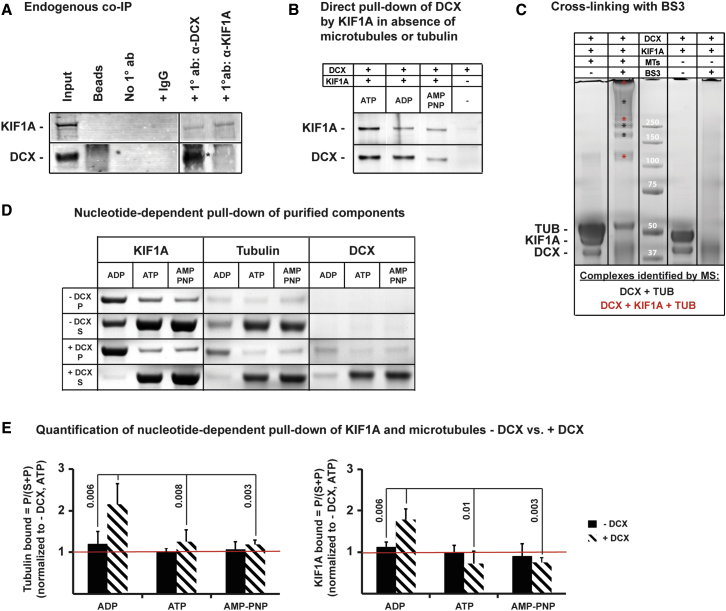
Doublecortin (Dcx) defines a growing family of microtubule (MT)-associated proteins (MAPs) involved in neuronal migration and process outgrowth. We show that Dcx is essential for the function of Kif1a, a kinesin-3 motor protein that traffics synaptic vesicles. Neurons lacking Dcx and/or its structurally conserved paralogue, doublecortin-like kinase 1 (Dclk1), show impaired Kif1a-mediated transport of Vamp2, a cargo of Kif1a, with decreased run length. Human disease-associated mutations in Dcx's linker sequence (e.g., W146C, K174E) alter Kif1a/Vamp2 transport by disrupting Dcx/Kif1a interactions without affecting Dcx MT binding. Dcx specifically enhances binding of the ADP-bound Kif1a motor domain to MTs. Cryo-electron microscopy and subnanometer-resolution image reconstruction reveal the kinesin-dependent conformational variability of MT-bound Dcx and suggest a model for MAP-motor crosstalk on MTs. Alteration of kinesin run length by MAPs represents a previously undiscovered mode of control of kinesin transport and provides a mechanism for regulation of MT-based transport by local signals.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures





References
-
- Bai J., Ramos R.L., Ackman J.B., Thomas A.M., Lee R.V., LoTurco J.J. RNAi reveals doublecortin is required for radial migration in rat neocortex. Nat. Neurosci. 2003;6:1277–1283. - PubMed
-
- Binder L.I., Frankfurter A., Rebhun L.I. Differential localization of MAP-2 and tau in mammalian neurons in situ. Ann. N Y Acad. Sci. 1986;466:145–166. - PubMed
-
- Caviston J.P., Holzbaur E.L. Microtubule motors at the intersection of trafficking and transport. Trends Cell Biol. 2006;16:530–537. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- WT_/Wellcome Trust/United Kingdom
- P01NS40043/NS/NINDS NIH HHS/United States
- R01AR055686/AR/NIAMS NIH HHS/United States
- 1F32HD070549-01/HD/NICHD NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P01 NS040043/NS/NINDS NIH HHS/United States
- MR/J000973/1/MRC_/Medical Research Council/United Kingdom
- P30 HD018655/HD/NICHD NIH HHS/United States
- R01 NS035129/NS/NINDS NIH HHS/United States
- T32 NS007473/NS/NINDS NIH HHS/United States
- K12 HD051959/HD/NICHD NIH HHS/United States
- T32 NS007484/NS/NINDS NIH HHS/United States
- 5R21NS063185-02/NS/NINDS NIH HHS/United States
- 2T32NS007473-11/NS/NINDS NIH HHS/United States
- F32 HD070549/HD/NICHD NIH HHS/United States
- R01 AR055686/AR/NIAMS NIH HHS/United States
- P30-HD-18655/HD/NICHD NIH HHS/United States
- K12HD051959-01/HD/NICHD NIH HHS/United States
- R21 NS063185/NS/NINDS NIH HHS/United States
- 2T32NS007484-11/NS/NINDS NIH HHS/United States
- R01NS35129/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
