

Plant-ants use symbiotic fungi as a food source: new insight into the nutritional ecology of ant-plant interactions
- PMID: 22859596
- PMCID: PMC3427587
- DOI: 10.1098/rspb.2012.1403
Plant-ants use symbiotic fungi as a food source: new insight into the nutritional ecology of ant-plant interactions
Abstract
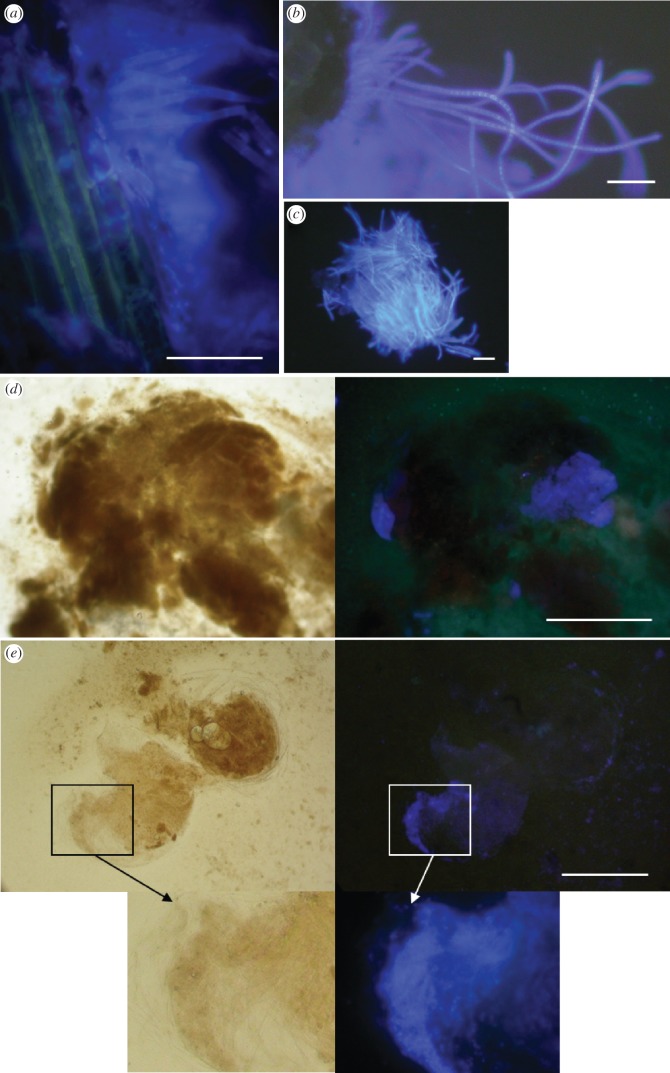
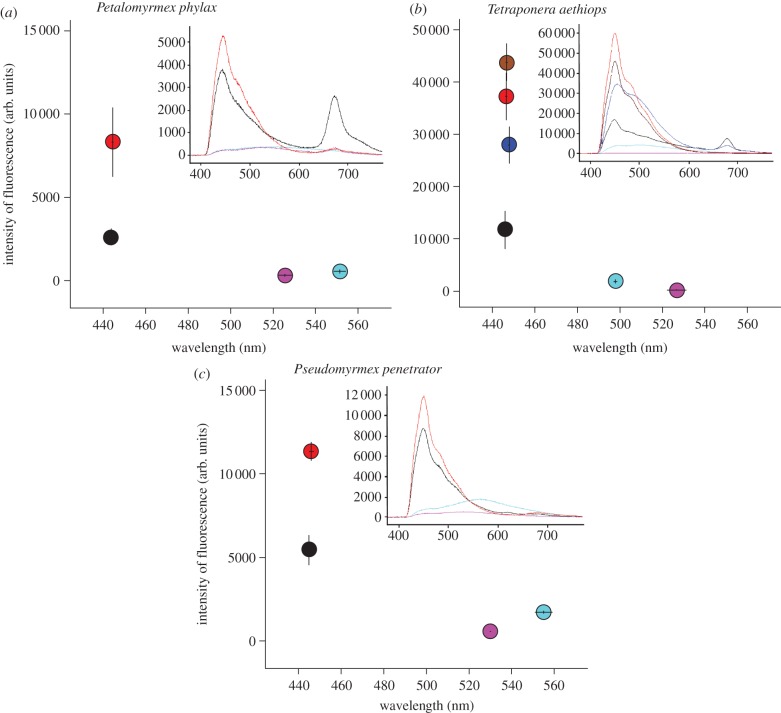
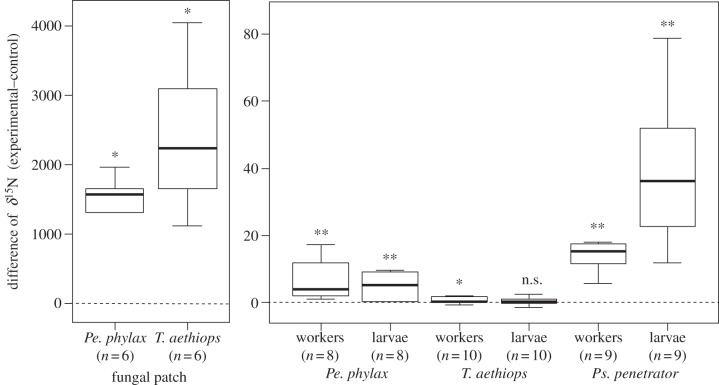
Usually studied as pairwise interactions, mutualisms often involve networks of interacting species. Numerous tropical arboreal ants are specialist inhabitants of myrmecophytes (plants bearing domatia, i.e. hollow structures specialized to host ants) and are thought to rely almost exclusively on resources derived from the host plant. Recent studies, following up on century-old reports, have shown that fungi of the ascomycete order Chaetothyriales live in symbiosis with plant-ants within domatia. We tested the hypothesis that ants use domatia-inhabiting fungi as food in three ant-plant symbioses: Petalomyrmex phylax/Leonardoxa africana, Tetraponera aethiops/Barteria fistulosa and Pseudomyrmex penetrator/Tachigali sp. Labelling domatia fungal patches in the field with either a fluorescent dye or (15)N showed that larvae ingested domatia fungi. Furthermore, when the natural fungal patch was replaced with a piece of a (15)N-labelled pure culture of either of two Chaetothyriales strains isolated from T. aethiops colonies, these fungi were also consumed. These two fungi often co-occur in the same ant colony. Interestingly, T. aethiops workers and larvae ingested preferentially one of the two strains. Our results add a new piece in the puzzle of the nutritional ecology of plant-ants.
Figures
References
-
- McKey D., Gaume L., Brouat C., Di Giusto B., Pascal L., Debout G., Dalecky A., Heil M. 2005. The trophic structure of tropical ant–plant–herbivore interactions: community consequences and coevolutionary dynamics. In Biotic interactions in the tropics: their role in the maintenance of species diversity (eds Burslem D., Pinard M., Hartley S.), pp. 386–413 Cambridge, UK: Cambridge University Press
-
- Heil M., McKey D. 2003. Protective ant–plant interactions as model systems in ecological and evolutionary research. Annu. Rev. Ecol. Evol. Syst. 34, 425–453 10.1146/annurev.ecolsys.34.011802.132410 (doi:10.1146/annurev.ecolsys.34.011802.132410) - DOI
-
- Davidson D. W., Cook S. C., Snelling R. R., Chua T. H. 2003. Explaining the abundance of ants in lowland tropical rainforest canopies. Science 300, 969–972 10.1126/science.1082074 (doi:10.1126/science.1082074) - DOI - PubMed
-
- Cook S. C., Davidson D. W. 2006. Nutritional and functional biology of exudate-feeding ants. Entomol. Exp. Appl. 118, 1–10 10.1111/j.1570-7458.2006.00374.x (doi:10.1111/j.1570-7458.2006.00374.x) - DOI
-
- Davidson D. W., McKey D. 1993. The evolutionary ecology of symbiotic ant–plant relationships. J. Hymenopt. Res. 2, 13–83
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources