

The natural history of class I primate alcohol dehydrogenases includes gene duplication, gene loss, and gene conversion
- PMID: 22859968
- PMCID: PMC3409193
- DOI: 10.1371/journal.pone.0041175
The natural history of class I primate alcohol dehydrogenases includes gene duplication, gene loss, and gene conversion
Abstract
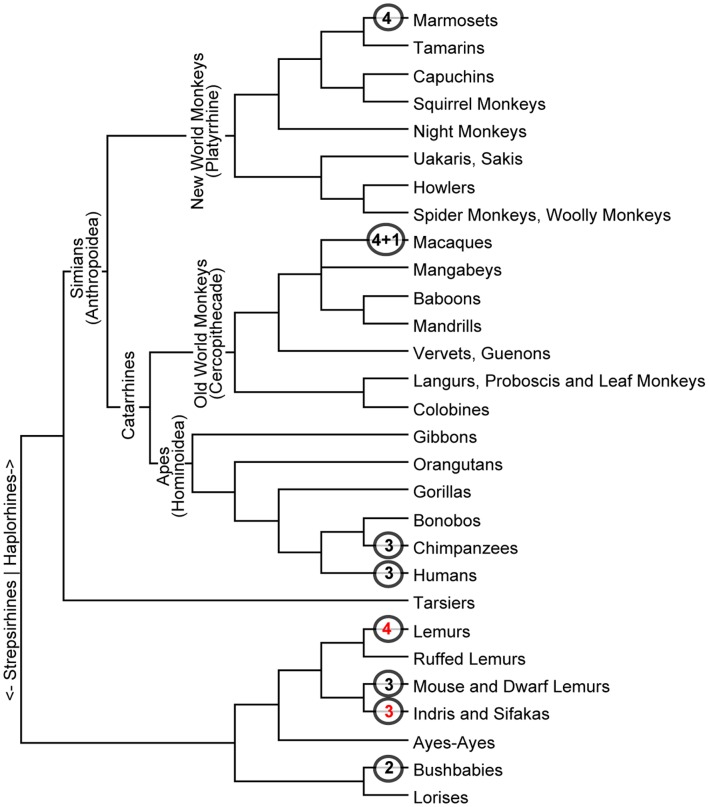
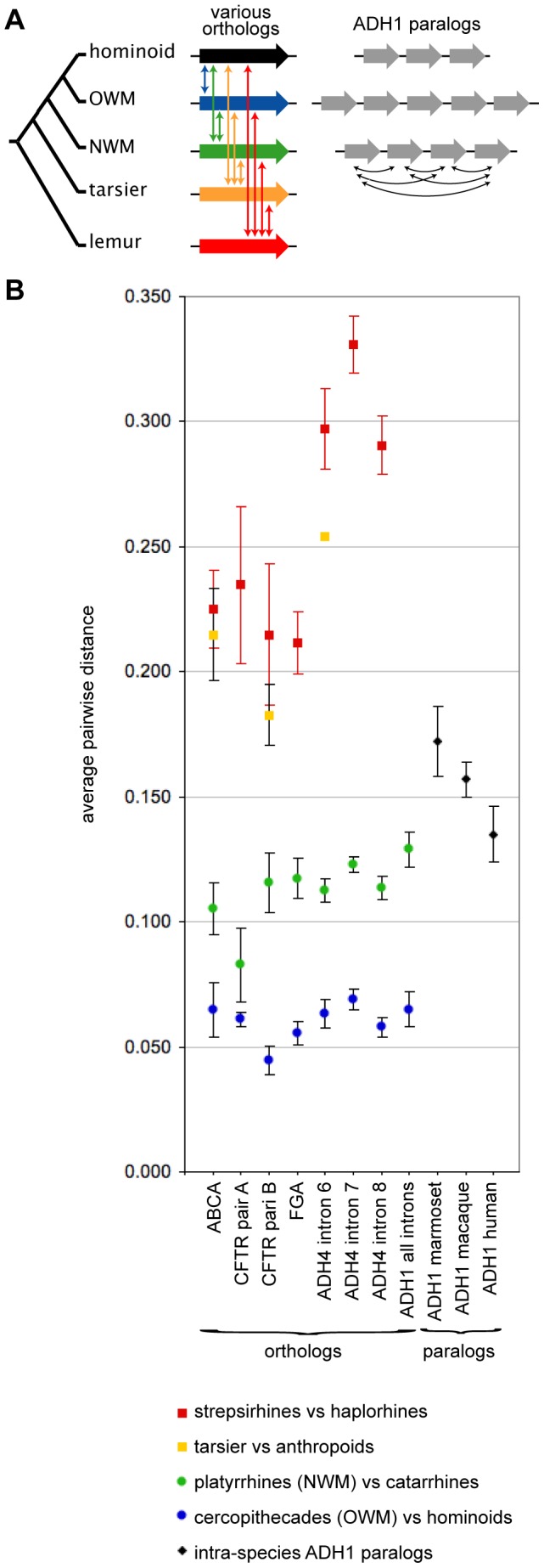
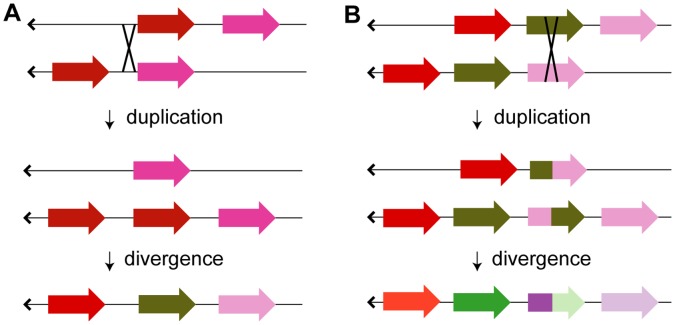
Background: Gene duplication is a source of molecular innovation throughout evolution. However, even with massive amounts of genome sequence data, correlating gene duplication with speciation and other events in natural history can be difficult. This is especially true in its most interesting cases, where rapid and multiple duplications are likely to reflect adaptation to rapidly changing environments and life styles. This may be so for Class I of alcohol dehydrogenases (ADH1s), where multiple duplications occurred in primate lineages in Old and New World monkeys (OWMs and NWMs) and hominoids.
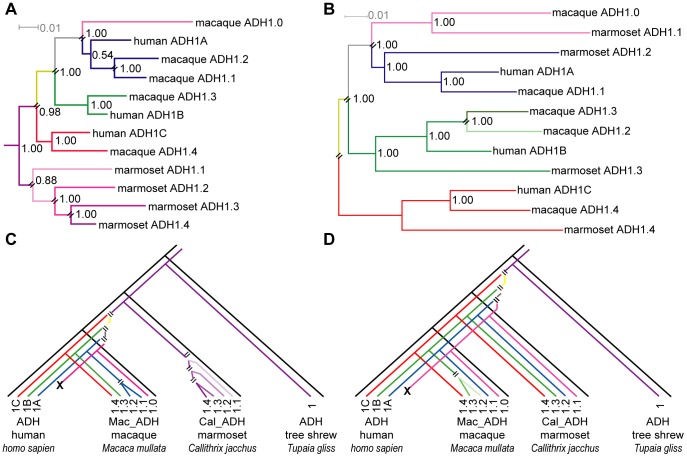
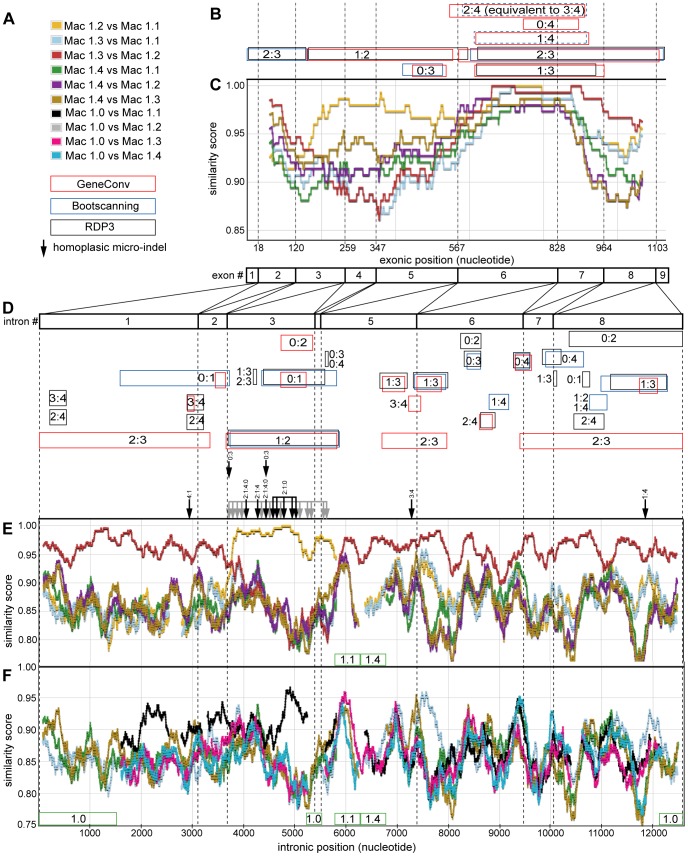
Methodology/principal findings: To build a preferred model for the natural history of ADH1s, we determined the sequences of nine new ADH1 genes, finding for the first time multiple paralogs in various prosimians (lemurs, strepsirhines). Database mining then identified novel ADH1 paralogs in both macaque (an OWM) and marmoset (a NWM). These were used with the previously identified human paralogs to resolve controversies relating to dates of duplication and gene conversion in the ADH1 family. Central to these controversies are differences in the topologies of trees generated from exonic (coding) sequences and intronic sequences.
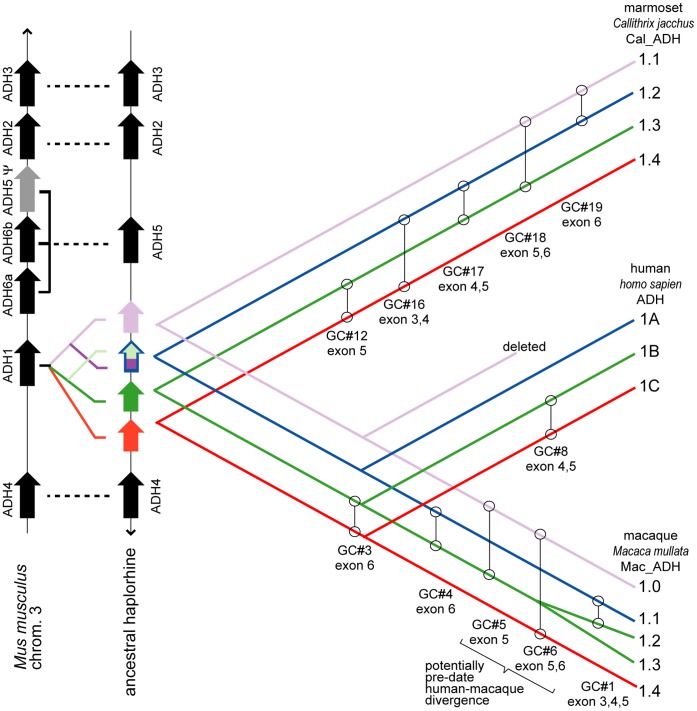
Conclusions/significance: We provide evidence that gene conversions are the primary source of difference, using molecular clock dating of duplications and analyses of microinsertions and deletions (micro-indels). The tree topology inferred from intron sequences appear to more correctly represent the natural history of ADH1s, with the ADH1 paralogs in platyrrhines (NWMs) and catarrhines (OWMs and hominoids) having arisen by duplications shortly predating the divergence of OWMs and NWMs. We also conclude that paralogs in lemurs arose independently. Finally, we identify errors in database interpretation as the source of controversies concerning gene conversion. These analyses provide a model for the natural history of ADH1s that posits four ADH1 paralogs in the ancestor of Catarrhine and Platyrrhine primates, followed by the loss of an ADH1 paralog in the human lineage.
Conflict of interest statement
Figures
References
-
- Bailey JA, Eichler EE (2006) Primate segmental duplications: crucibles of evolution, diversity and disease. Nature Reviews Genetics 7: 552–564. - PubMed
-
- Chang M, Benner SA (2004) Empirical analysis of insertions and deletions in protein sequence evolution. J Mol Biol 341: 617–631. - PubMed
-
- Fukami-Kobayashi K, Schreiber DR, Benner SA (2002) Detecting compensatory covariation signals in protein evolution using reconstructed ancestral sequences. J Mol Biol 319: 729–743. - PubMed
-
- Sassi SO, Braun EL, Benner SA (2007) The evolution of seminal ribonuclease. Pseudogene reactivation or multiple gene inactivation events? Mol Biol Evol 24: 1012–1024. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
