

Proteomic changes in rat spermatogenesis in response to in vivo androgen manipulation; impact on meiotic cells
- PMID: 22860010
- PMCID: PMC3408499
- DOI: 10.1371/journal.pone.0041718
Proteomic changes in rat spermatogenesis in response to in vivo androgen manipulation; impact on meiotic cells
Abstract
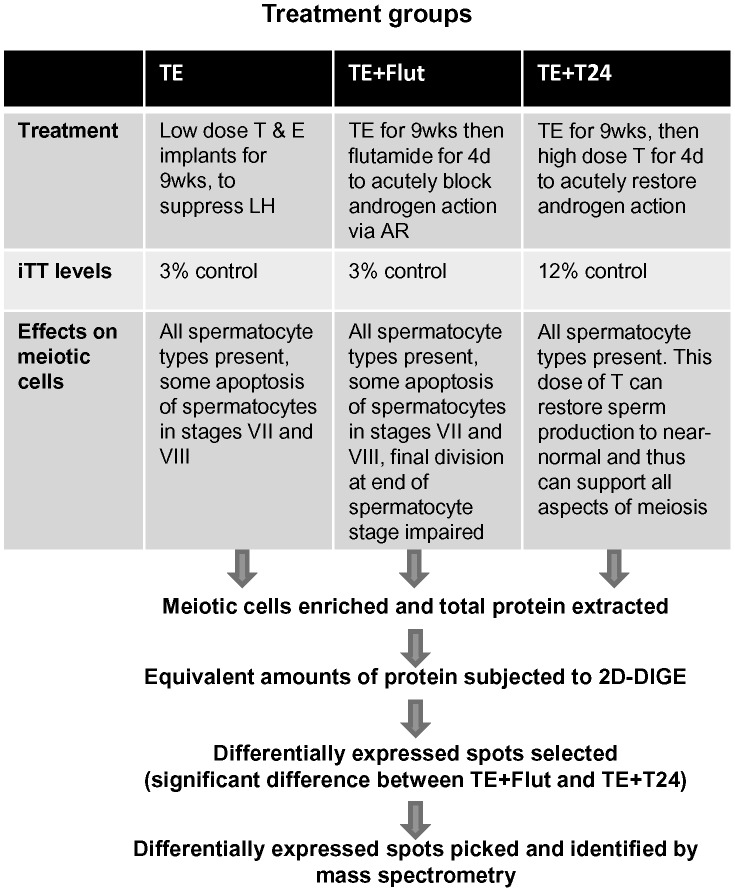
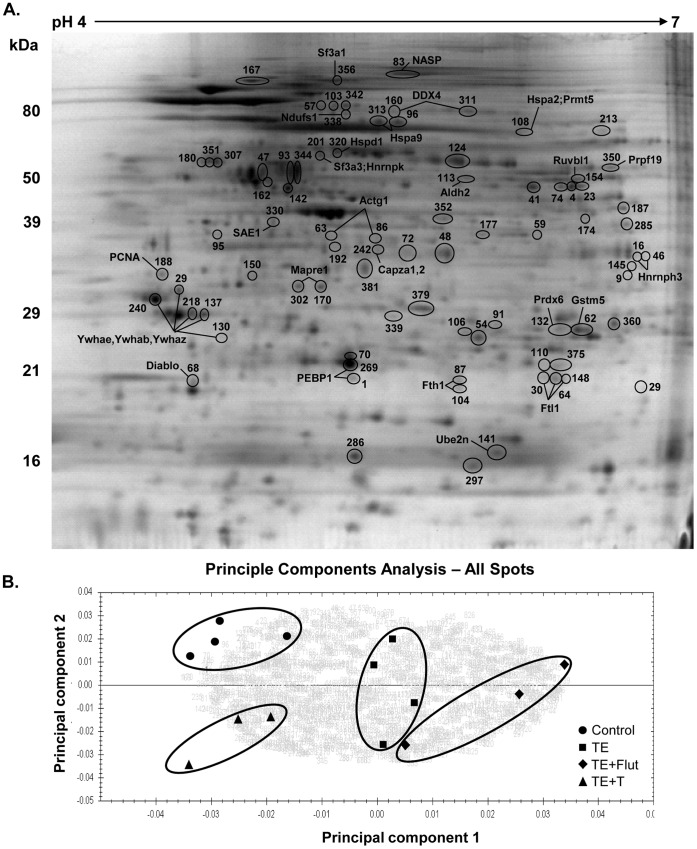
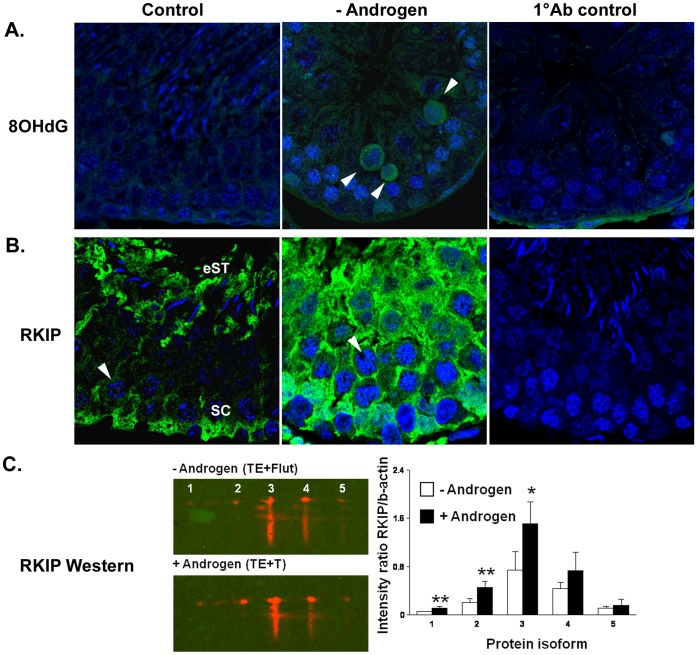
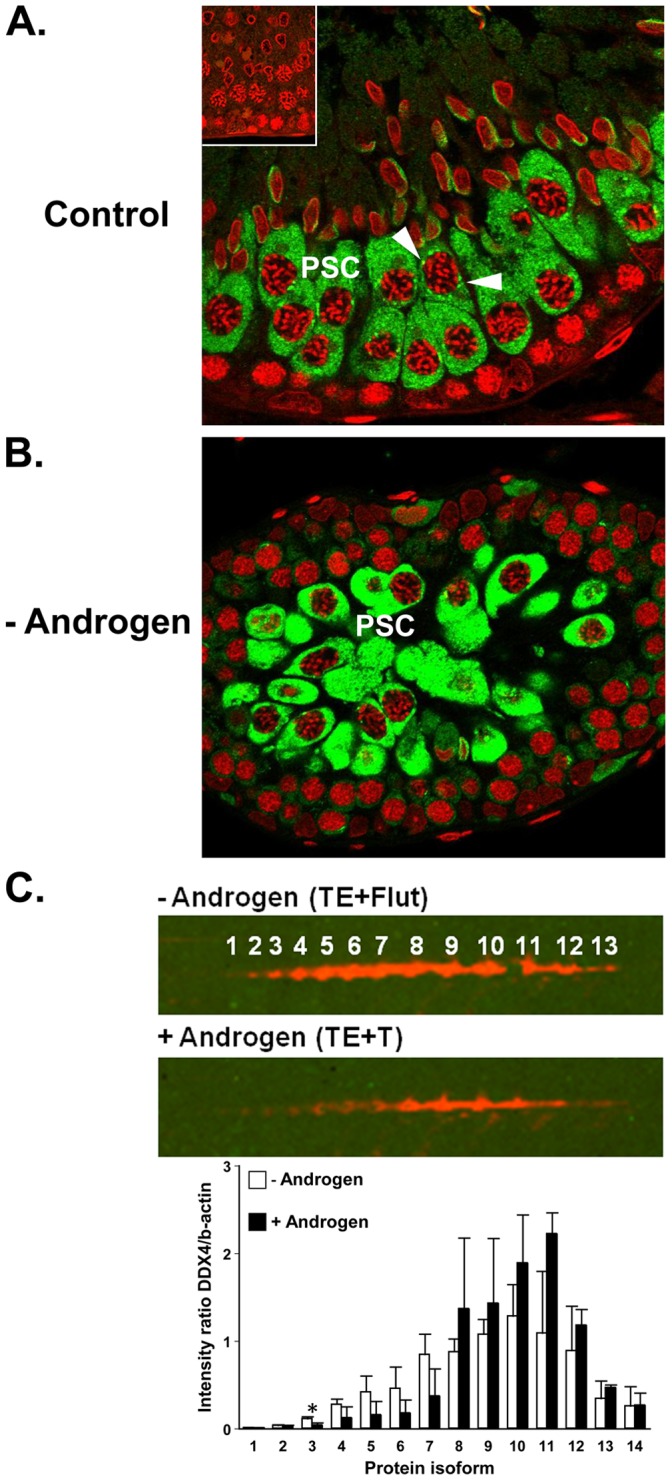
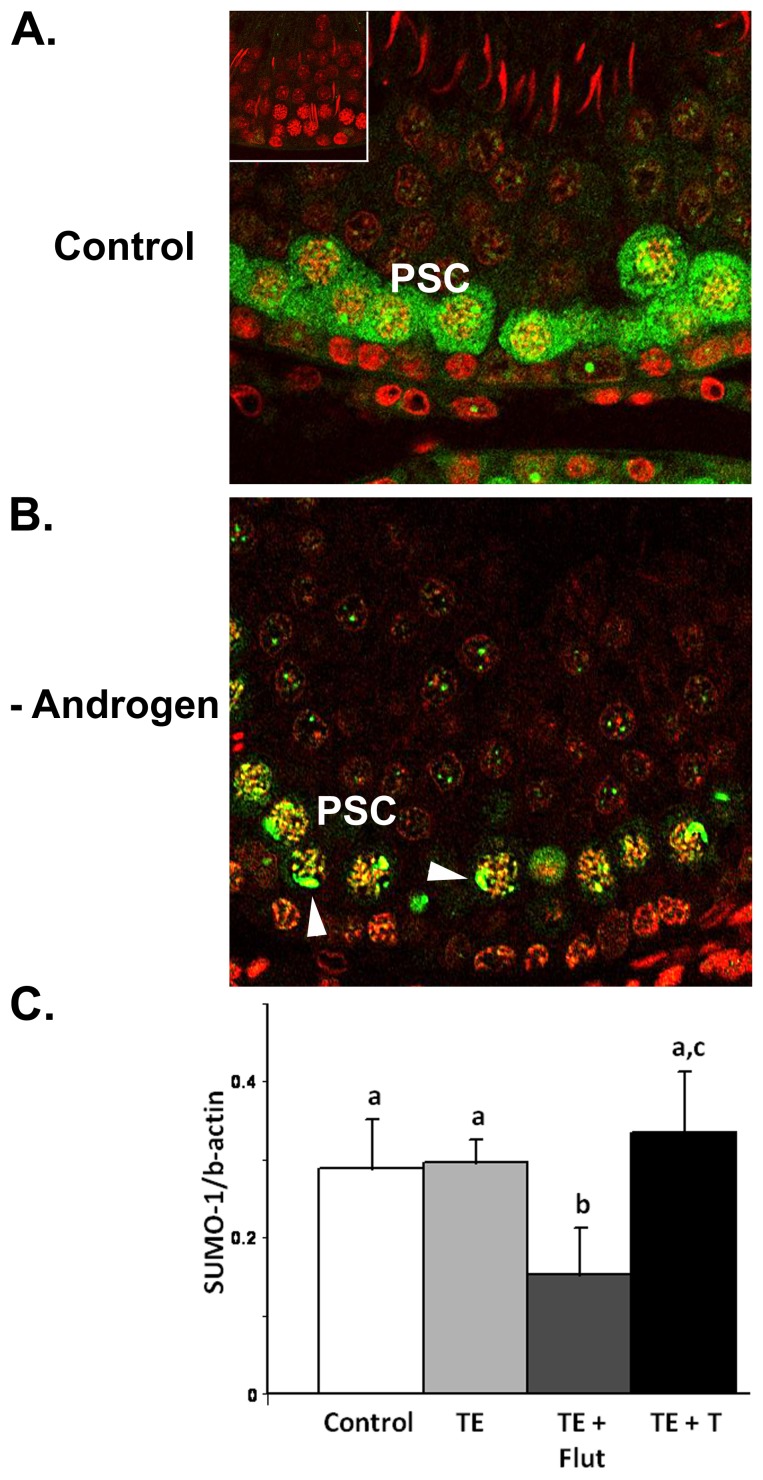
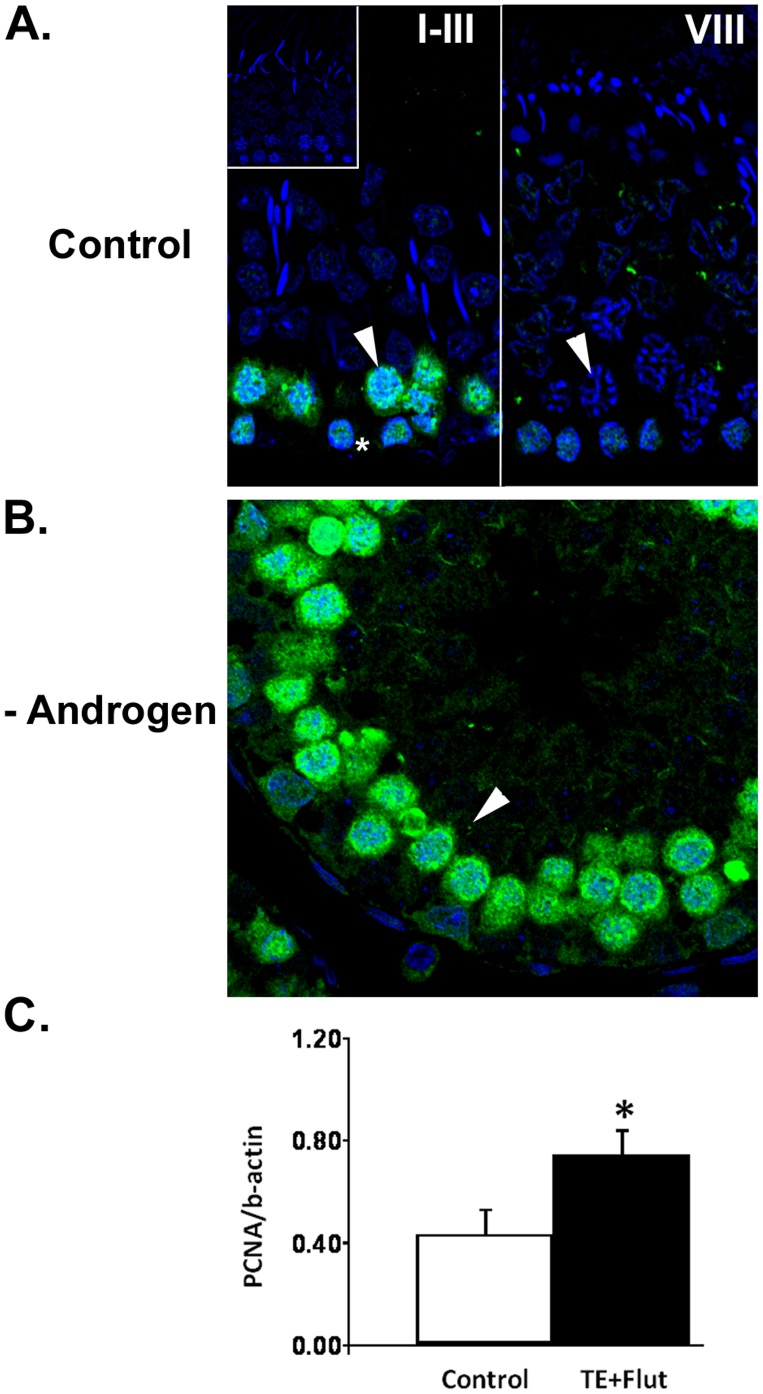
The production of mature sperm is reliant on androgen action within the testis, and it is well established that androgens act on receptors within the somatic Sertoli cells to stimulate male germ cell development. Mice lacking Sertoli cell androgen receptors (AR) show late meiotic germ cell arrest, suggesting Sertoli cells transduce the androgenic stimulus co-ordinating this essential step in spermatogenesis. This study aimed to identify germ cell proteins responsive to changes in testicular androgen levels and thereby elucidate mechanisms by which androgens regulate meiosis. Testicular androgen levels were suppressed for 9 weeks using testosterone and estradiol-filled silastic implants, followed by a short period of either further androgen suppression (via an AR antagonist) or the restoration of intratesticular testosterone levels. Comparative proteomics were performed on protein extracts from enriched meiotic cell preparations from adult rats undergoing androgen deprivation and replacement in vivo. Loss of androgenic stimulus caused changes in proteins with known roles in meiosis (including Nasp and Hsp70-2), apoptosis (including Diablo), cell signalling (including 14-3-3 isoforms), oxidative stress, DNA repair, and RNA processing. Immunostaining for oxidised DNA adducts confirmed spermatocytes undergo oxidative stress-induced DNA damage during androgen suppression. An increase in PCNA and an associated ubiquitin-conjugating enzyme (Ubc13) suggested a role for PCNA-mediated regulation of DNA repair pathways in spermatocytes. Changes in cytoplasmic SUMO1 localisation in spermatocytes were paralleled by changes in the levels of free SUMO1 and of a subunit of its activating complex, suggesting sumoylation in spermatocytes is modified by androgen action on Sertoli cells. We conclude that Sertoli cells, in response to androgens, modulate protein translation and post-translational events in spermatocytes that impact on their metabolism, survival, and completion of meiosis.
Conflict of interest statement
Figures
Similar articles
-
Intratesticular androgen levels, androgen receptor localization, and androgen receptor expression in adult rat Sertoli cells.Biol Reprod. 2004 Oct;71(4):1348-58. doi: 10.1095/biolreprod.104.029249. Epub 2004 Jun 23. Biol Reprod. 2004. PMID: 15215201
-
Androgen action on the restoration of spermatogenesis in adult rats: effects of human chorionic gonadotrophin, testosterone and flutamide administration on germ cell number.Int J Androl. 1997 Apr;20(2):70-9. doi: 10.1046/j.1365-2605.1997.d01-121.x. Int J Androl. 1997. PMID: 9292316
-
Testicular expression of small ubiquitin-related modifier-1 (SUMO-1) supports multiple roles in spermatogenesis: silencing of sex chromosomes in spermatocytes, spermatid microtubule nucleation, and nuclear reshaping.Dev Biol. 2005 Jun 15;282(2):480-92. doi: 10.1016/j.ydbio.2005.03.034. Dev Biol. 2005. PMID: 15950612
-
Regulation of seminiferous tubular function by FSH and androgen.J Reprod Fertil. 1975 Aug;44(2):363-75. doi: 10.1530/jrf.0.0440363. J Reprod Fertil. 1975. PMID: 169349 Review.
-
The regulation of spermatogenesis by androgens.Semin Cell Dev Biol. 2014 Jun;30:2-13. doi: 10.1016/j.semcdb.2014.02.012. Epub 2014 Mar 2. Semin Cell Dev Biol. 2014. PMID: 24598768 Free PMC article. Review.
Cited by
-
The Role of Testosterone in Spermatogenesis: Lessons From Proteome Profiling of Human Spermatozoa in Testosterone Deficiency.Front Endocrinol (Lausanne). 2022 May 19;13:852661. doi: 10.3389/fendo.2022.852661. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 35663320 Free PMC article.
-
Ubc13: the Lys63 ubiquitin chain building machine.Oncotarget. 2016 Sep 27;7(39):64471-64504. doi: 10.18632/oncotarget.10948. Oncotarget. 2016. PMID: 27486774 Free PMC article. Review.
-
Proteomic analysis of human spermatozoa proteins with oxidative stress.Reprod Biol Endocrinol. 2013 May 20;11:48. doi: 10.1186/1477-7827-11-48. Reprod Biol Endocrinol. 2013. PMID: 23688036 Free PMC article.
-
Quantitative SWATH-Based Proteomic Profiling for Identification of Mechanism-Driven Diagnostic Biomarkers Conferring in the Progression of Metastatic Prostate Cancer.Front Oncol. 2020 Apr 8;10:493. doi: 10.3389/fonc.2020.00493. eCollection 2020. Front Oncol. 2020. PMID: 32322560 Free PMC article.
-
RKIP suppresses the proliferation and invasion ofchoriocarcinoma cells through inhibiting the MAPK signaling pathway.Int J Clin Exp Med. 2015 Dec 15;8(12):22183-90. eCollection 2015. Int J Clin Exp Med. 2015. PMID: 26885193 Free PMC article.
References
-
- Petersen C, Soder O (2006) The Sertoli cell–a hormonal target and 'super' nurse for germ cells that determines testicular size. Horm Res 66: 153–161. - PubMed
-
- O'Donnell L, Meachem SJ, Stanton PG, McLachlan RI (2006) Endocrine regulation of spermatogenesis. In: Neill JD, editor. Knobil and Neill's Physiology of Reproduction. 3rd ed. San Diego, CA: Elsevier. 1017–1069.
-
- Walker WH, Cheng J (2005) FSH and testosterone signaling in Sertoli cells. Reproduction 130: 15–28. - PubMed
-
- Ruwanpura SM, McLachlan RI, Meachem SJ (2010) Hormonal regulation of male germ cell development. J Endocrinol 205: 117–131. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
