

Characterization of Rad51 from apicomplexan parasite Toxoplasma gondii: an implication for inefficient gene targeting
- PMID: 22860032
- PMCID: PMC3408395
- DOI: 10.1371/journal.pone.0041925
Characterization of Rad51 from apicomplexan parasite Toxoplasma gondii: an implication for inefficient gene targeting
Abstract
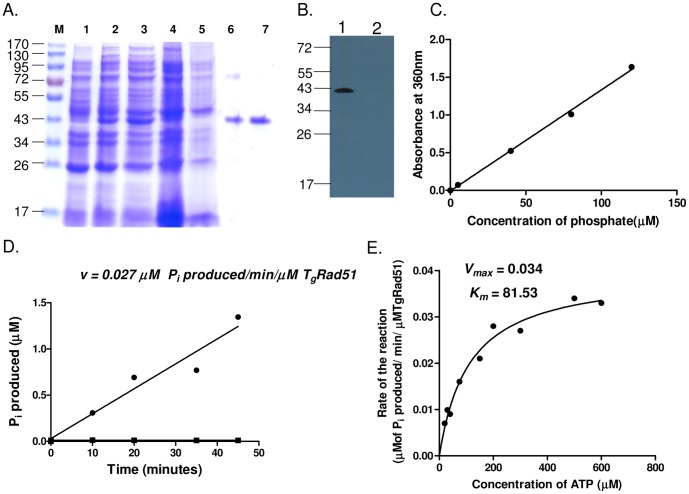
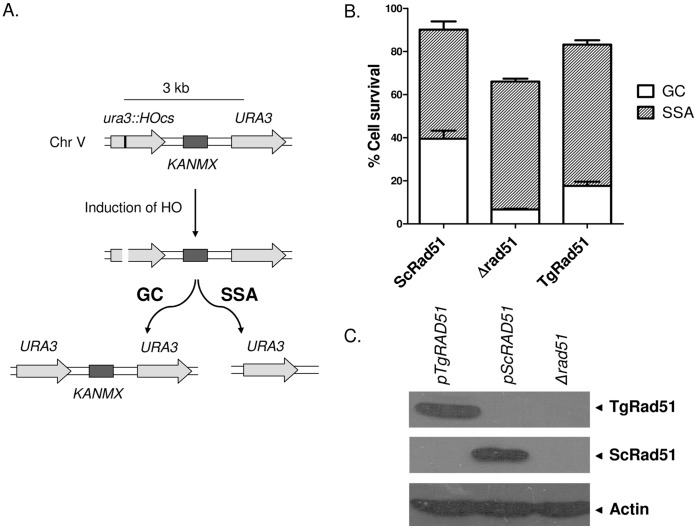
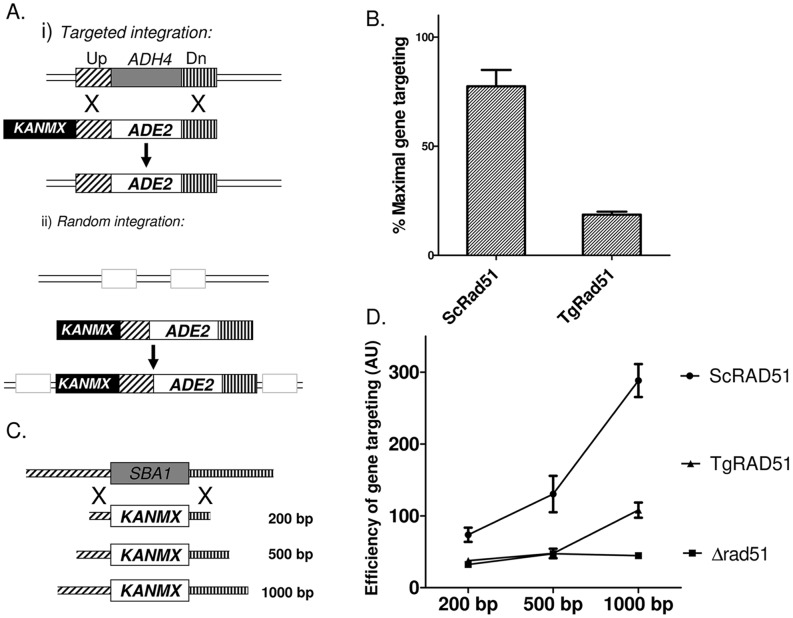
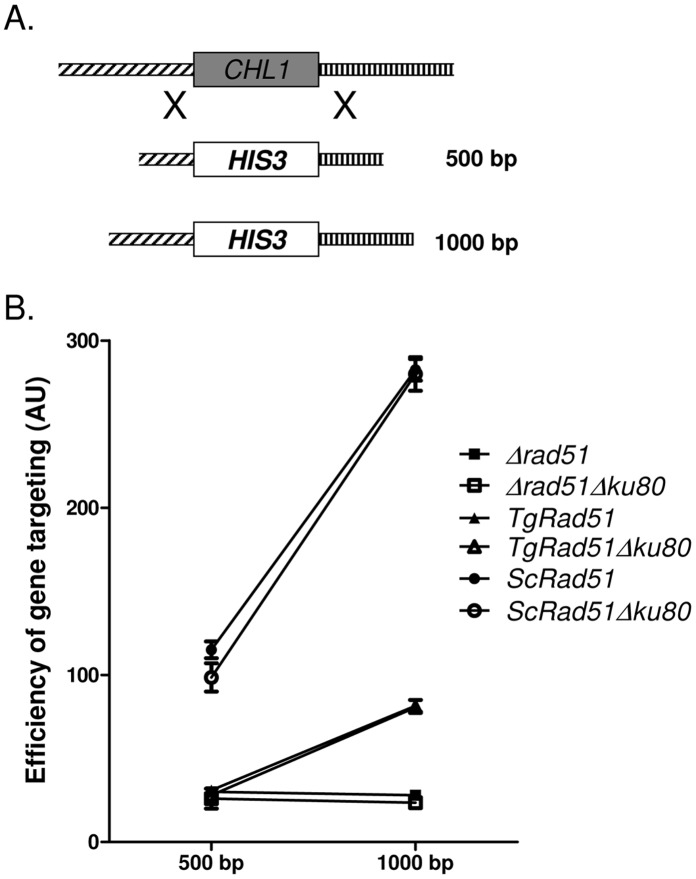
Repairing double strand breaks (DSBs) is absolutely essential for the survival of obligate intracellular parasite Toxoplasma gondii. Thus, DSB repair mechanisms could be excellent targets for chemotherapeutic interventions. Recent genetic and bioinformatics analyses confirm the presence of both homologous recombination (HR) as well as non homologous end joining (NHEJ) proteins in this lower eukaryote. In order to get mechanistic insights into the HR mediated DSB repair pathway in this parasite, we have characterized the key protein involved in homologous recombination, namely TgRad51, at the biochemical and genetic levels. We have purified recombinant TgRad51 protein to 99% homogeneity and have characterized it biochemically. The ATP hydrolysis activity of TgRad51 shows a higher K(M) and much lower k(cat) compared to bacterial RecA or Rad51 from other related protozoan parasites. Taking yeast as a surrogate model system we have shown that TgRad51 is less efficient in gene conversion mechanism. Further, we have found that TgRad51 mediated gene integration is more prone towards random genetic loci rather than targeted locus. We hypothesize that compromised ATPase activity of TgRad51 is responsible for inefficient gene targeting and poor gene conversion efficiency in this protozoan parasite. With increase in homologous flanking regions almost three fold increments in targeted gene integration is observed, which is similar to the trend found with ScRad51. Our findings not only help us in understanding the reason behind inefficient gene targeting in T. gondii but also could be exploited to facilitate high throughput knockout as well as epitope tagging of Toxoplasma genes.
Conflict of interest statement
Figures

References
-
- Haber JE (2000) Partners and pathways repairing a double strand break. Trends in Genetics 16(6): 259–264. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
