

Sizzled-tolloid interactions maintain foregut progenitors by regulating fibronectin-dependent BMP signaling
- PMID: 22863744
- PMCID: PMC3615545
- DOI: 10.1016/j.devcel.2012.07.002
Sizzled-tolloid interactions maintain foregut progenitors by regulating fibronectin-dependent BMP signaling
Abstract
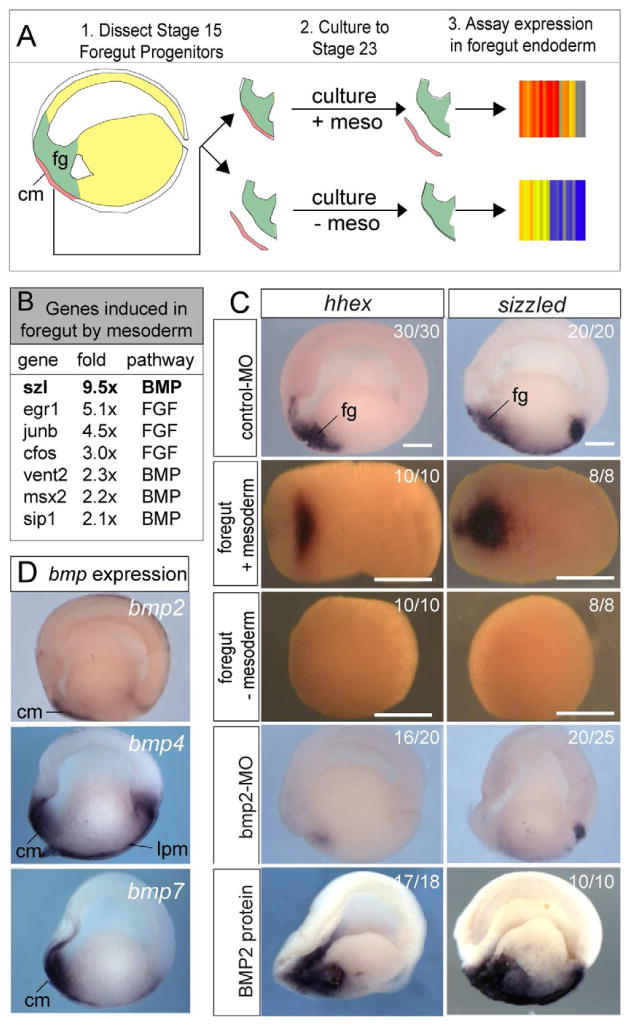
The liver, pancreas, and lungs are induced from endoderm progenitors by a series of dynamic growth factor signals from the mesoderm, but how the temporal-spatial activity of these signals is controlled is poorly understood. We have identified an extracellular regulatory loop required for robust bone morphogenetic protein (BMP) signaling in the Xenopus foregut. We show that BMP signaling is required to maintain foregut progenitors and induce expression of the secreted frizzled related protein Sizzled (Szl) and the extracellular metalloprotease Tolloid-like 1 (Tll1). Szl negatively regulates Tll activity to control deposition of a fibronectin (FN) matrix between the mesoderm and endoderm, which is required to maintain BMP signaling. Foregut-specific Szl depletion results in a loss of the FN matrix and failure to maintain robust pSmad1 levels, causing a loss of foregut gene expression and organ agenesis. These results have implications for BMP signaling in diverse contexts and the differentiation of foregut tissue from stem cells.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures







References
-
- Blitz IL, Shimmi O, Wunnenberg-Stapleton K, O’Connor MB, Cho KW. Is chordin a long-range- or short-range-acting factor? Roles for BMP1-related metalloproteases in chordin and BMP4 autofeedback loop regulation. Dev Biol. 2000;223:120–138. - PubMed
-
- Bovolenta P, Esteve P, Ruiz JM, Cisneros E, Lopez-Rios J. Beyond Wnt inhibition: new functions of secreted Frizzled-related proteins in development and disease. J Cell Sci. 2008;121:737–746. - PubMed
-
- Collavin L, Kirschner MW. The secreted Frizzled-related protein Sizzled functions as a negative feedback regulator of extreme ventral mesoderm. Development. 2003;130:805–816. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
