

Engineering a cyanobacterial cell factory for production of lactic acid
- PMID: 22865063
- PMCID: PMC3457509
- DOI: 10.1128/AEM.01587-12
Engineering a cyanobacterial cell factory for production of lactic acid
Abstract
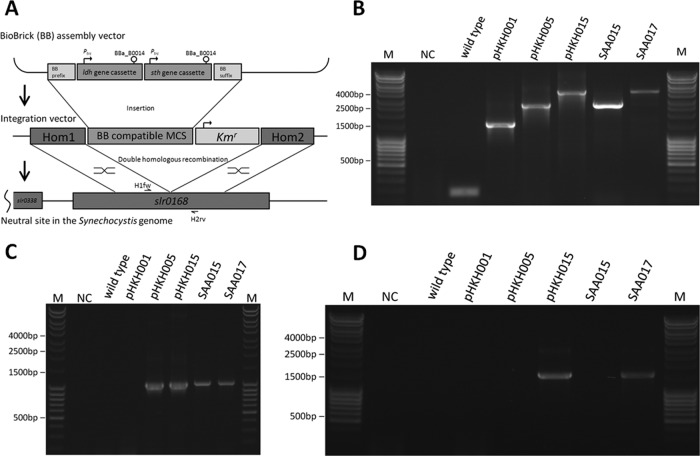
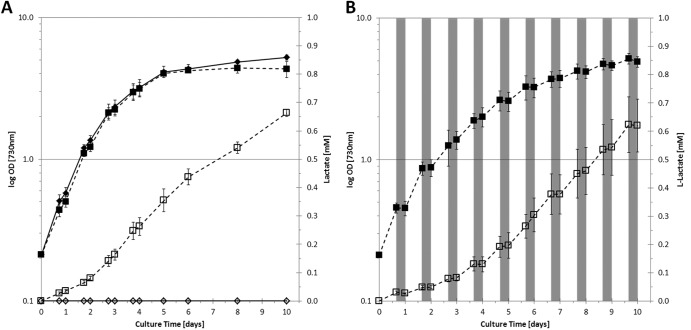
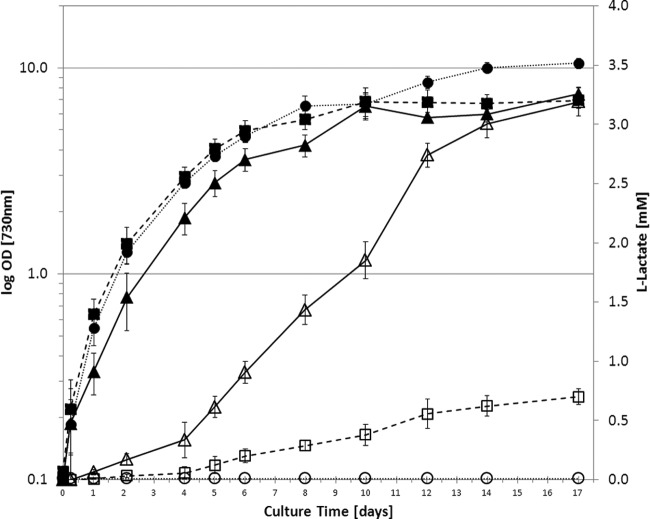
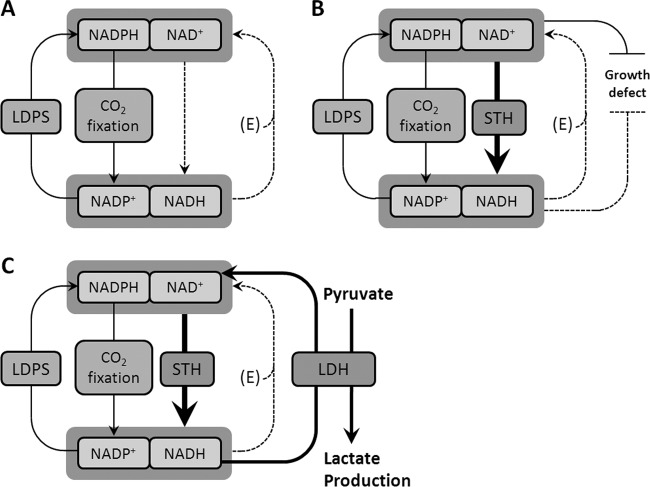
Metabolic engineering of microorganisms has become a versatile tool to facilitate production of bulk chemicals, fuels, etc. Accordingly, CO(2) has been exploited via cyanobacterial metabolism as a sustainable carbon source of biofuel and bioplastic precursors. Here we extended these observations by showing that integration of an ldh gene from Bacillus subtilis (encoding an l-lactate dehydrogenase) into the genome of Synechocystis sp. strain PCC6803 leads to l-lactic acid production, a phenotype which is shown to be stable for prolonged batch culturing. Coexpression of a heterologous soluble transhydrogenase leads to an even higher lactate production rate and yield (lactic acid accumulating up to a several-millimolar concentration in the extracellular medium) than those for the single ldh mutant. The expression of a transhydrogenase alone, however, appears to be harmful to the cells, and a mutant carrying such a gene is rapidly outcompeted by a revertant(s) with a wild-type growth phenotype. Furthermore, our results indicate that the introduction of a lactate dehydrogenase rescues this phenotype by preventing the reversion.
Figures
References
-
- Abdel-Rahman MA, Tashiro Y, Sonomoto K. 2010. Lactic acid production from lignocellulose-derived sugars using lactic acid bacteria: overview and limits. J. Biotechnol. 156: 286– 301 - PubMed
-
- Angermayr SA, Hellingwerf KJ, Lindblad P, de Mattos MJT. 2009. Energy biotechnology with cyanobacteria. Curr. Opin. Biotechnol. 20: 257– 263 - PubMed
-
- Atsumi S, Higashide W, Liao JC. 2009. Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nat. Biotechnol. 27: 1177– 1180 - PubMed
-
- Bentley FK, Melis A. 2012. Diffusion-based process for carbon dioxide uptake and isoprene emission in gaseous/aqueous two-phase photobioreactors by photosynthetic microorganisms. Biotechnol. Bioeng. 109: 100– 109 - PubMed
-
- Brosius J, Erfle M, Storella J. 1985. Spacing of the −10 and −35 regions in the tac promoter. Effect on its in vivo activity. J. Biol. Chem. 260: 3539– 3541 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
