

Dual lineage-specific expression of Sox17 during mouse embryogenesis
- PMID: 22865702
- PMCID: PMC3448801
- DOI: 10.1002/stem.1192
Dual lineage-specific expression of Sox17 during mouse embryogenesis
Abstract
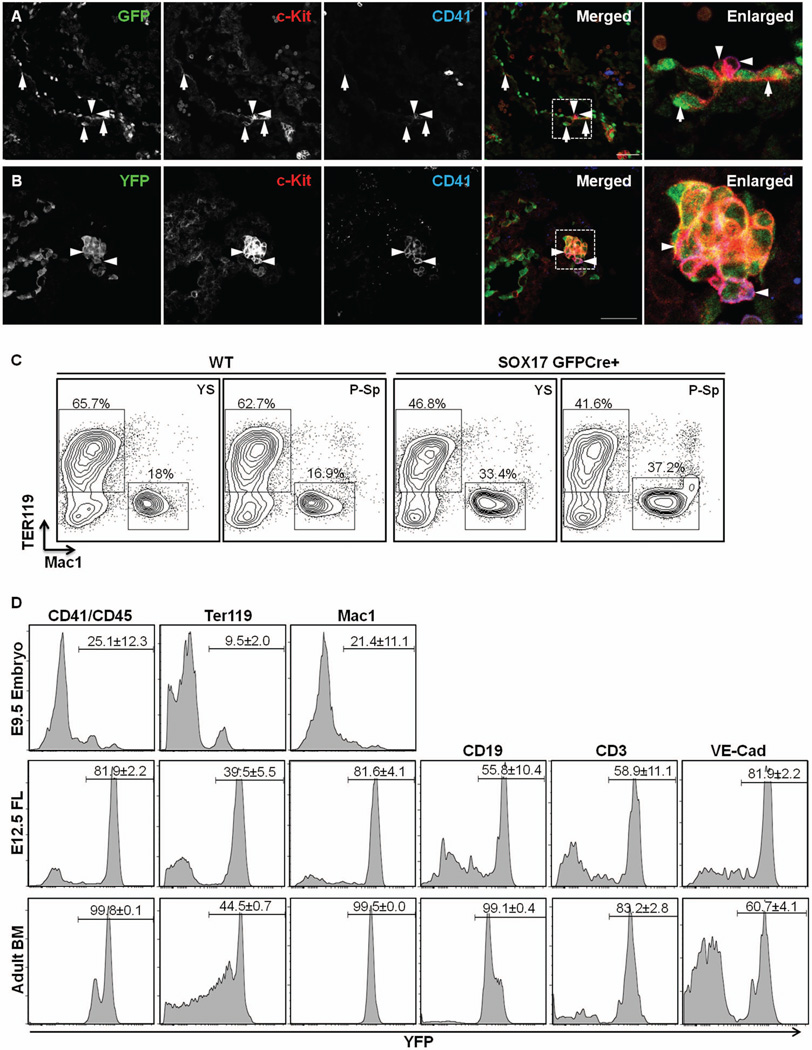
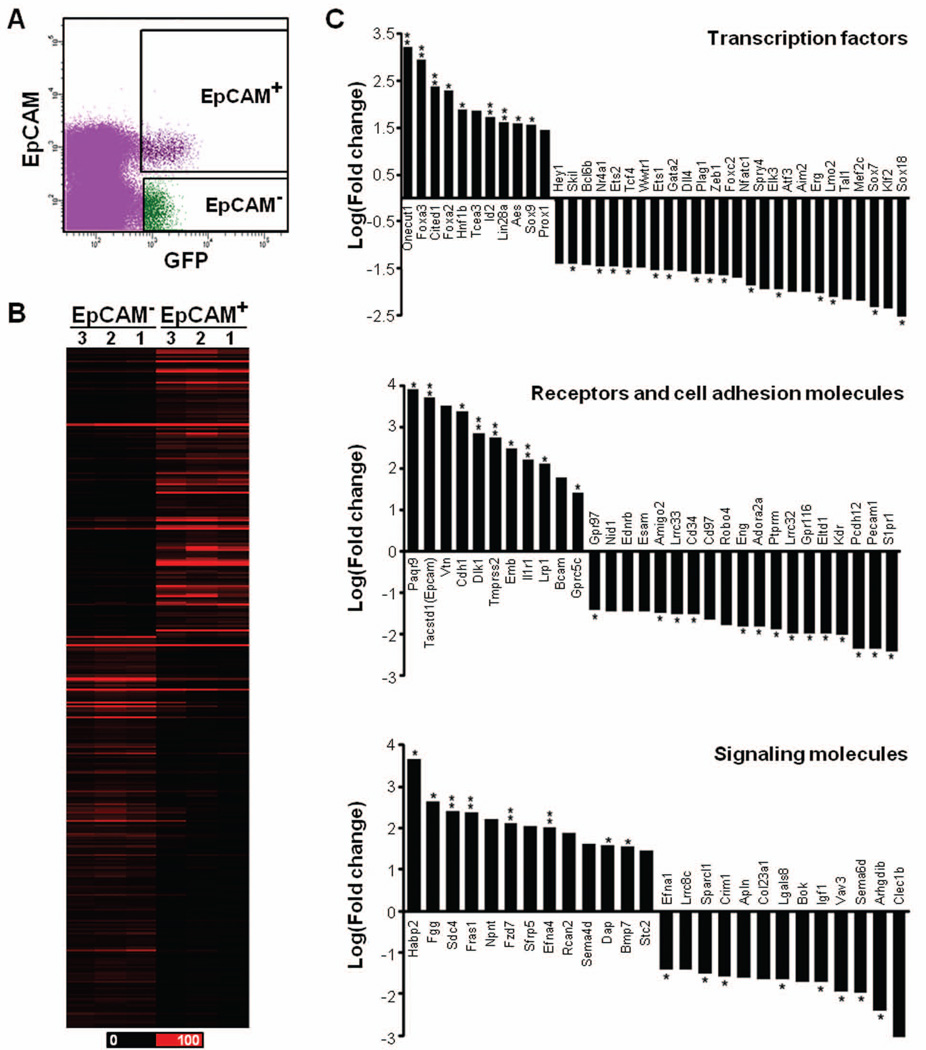
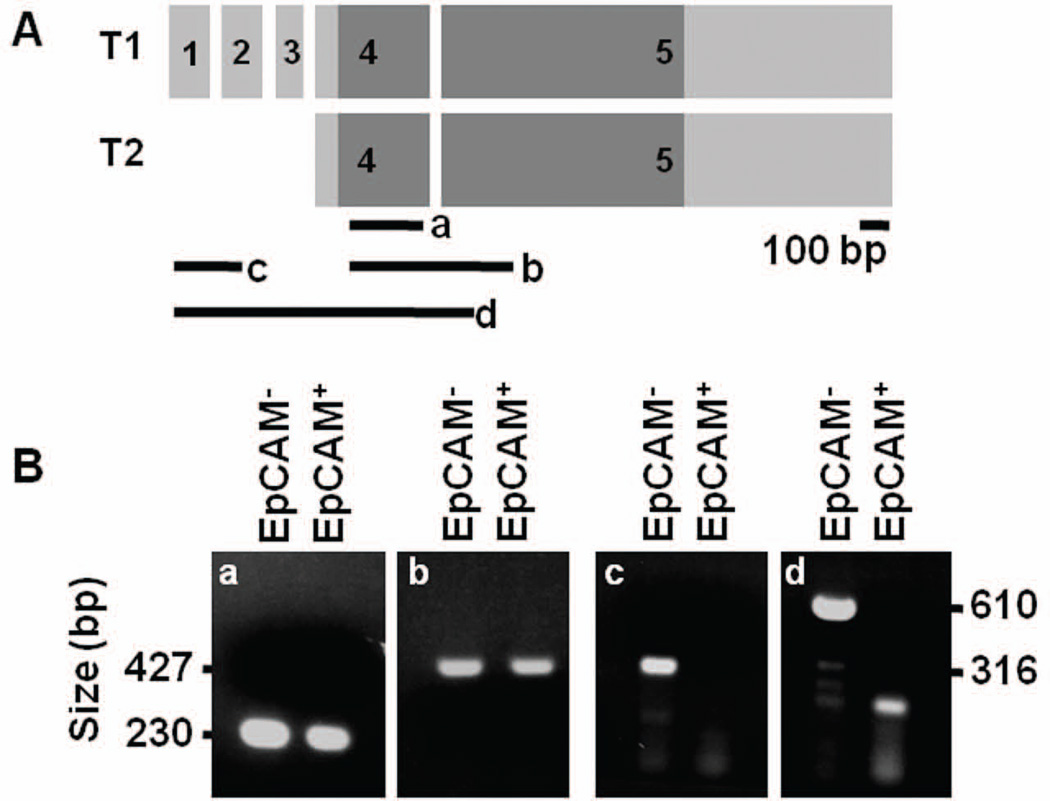
Sox17 is essential for both endoderm development and fetal hematopoietic stem cell (HSC) maintenance. While endoderm-derived organs are well known to originate from Sox17-expressing cells, it is less certain whether fetal HSCs also originate from Sox17-expressing cells. By generating a Sox17(GFPCre) allele and using it to assess the fate of Sox17-expressing cells during embryogenesis, we confirmed that both endodermal and a part of definitive hematopoietic cells are derived from Sox17-positive cells. Prior to E9.5, the expression of Sox17 is restricted to the endoderm lineage. However, at E9.5 Sox17 is expressed in the endothelial cells (ECs) at the para-aortic splanchnopleural region that contribute to the formation of HSCs at a later stage. The identification of two distinct progenitor cell populations that express Sox17 at E9.5 was confirmed using fluorescence-activated cell sorting together with RNA-Seq to determine the gene expression profiles of the two cell populations. Interestingly, this analysis revealed differences in the RNA processing of the Sox17 mRNA during embryogenesis. Taken together, these results indicate that Sox17 is expressed in progenitor cells derived from two different germ layers, further demonstrating the complex expression pattern of this gene and suggesting caution when using Sox17 as a lineage-specific marker.
Copyright © 2012 AlphaMed Press.
Conflict of interest statement
No potential conflicts of interest.
Figures
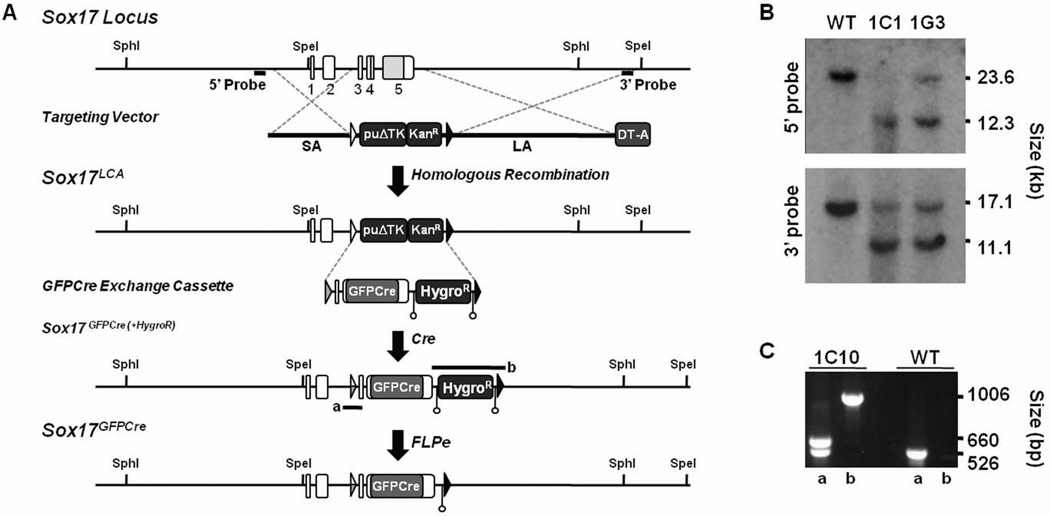
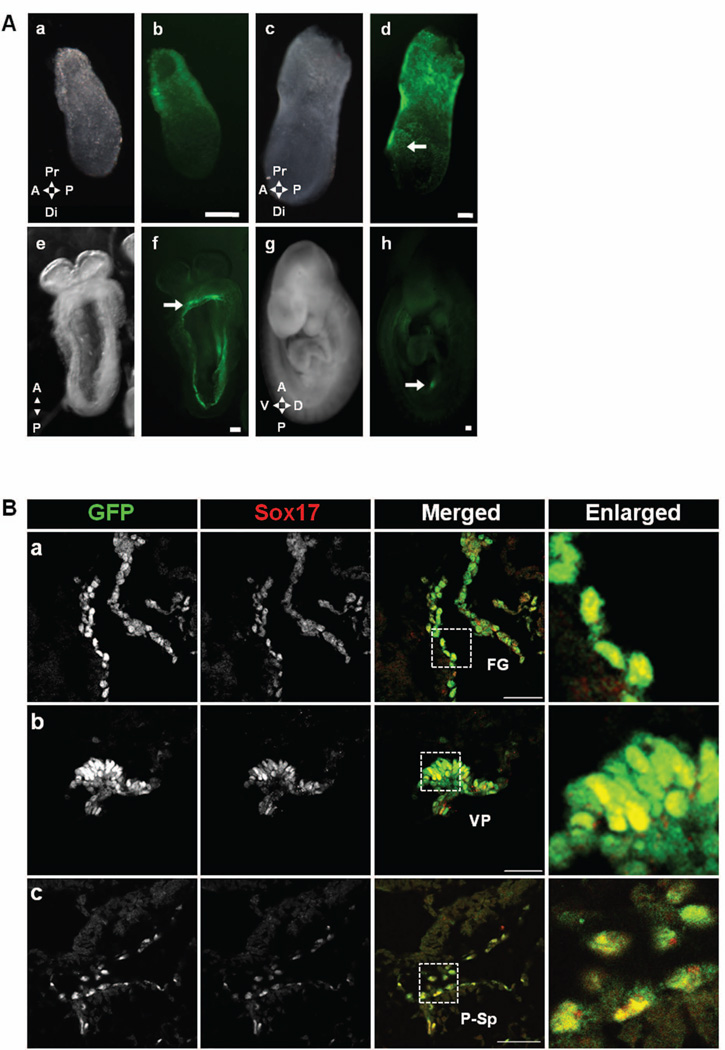
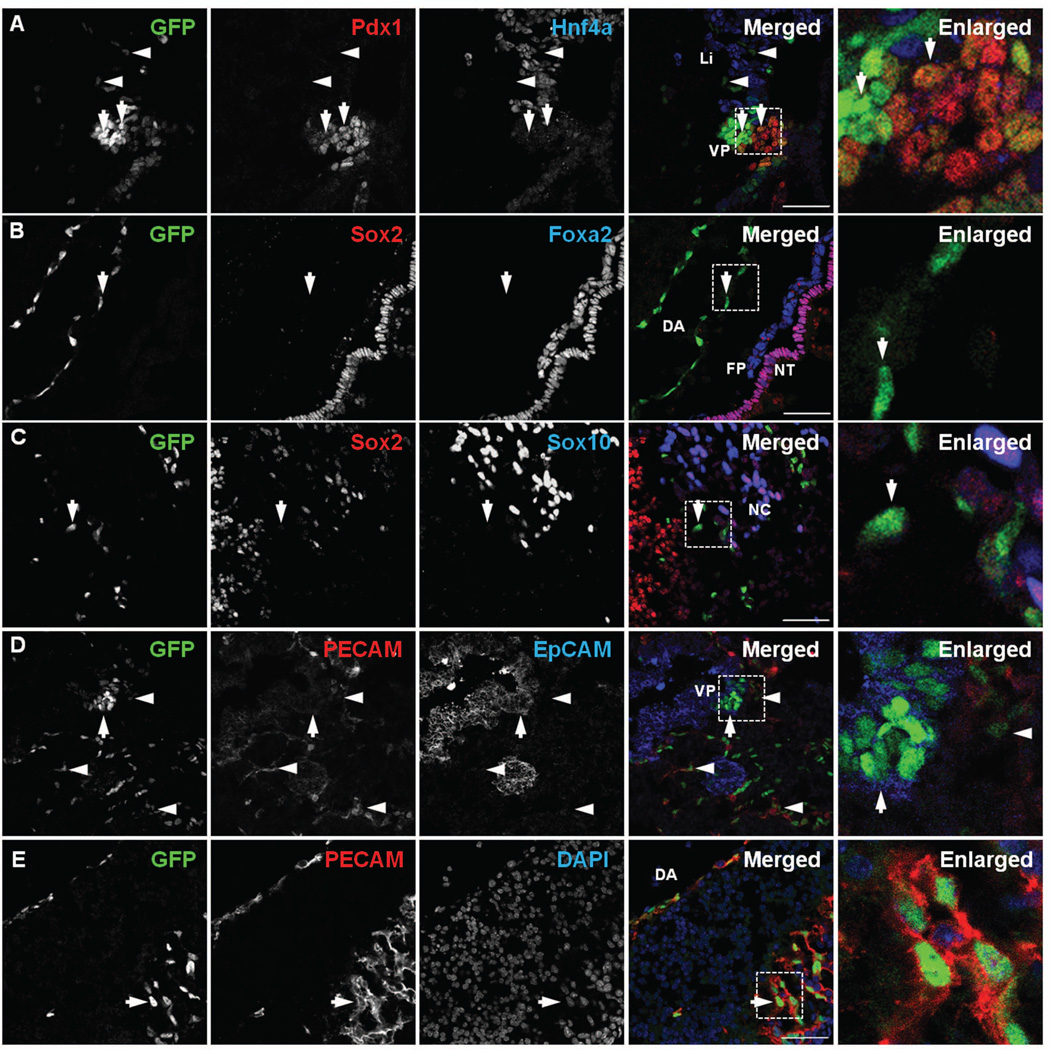
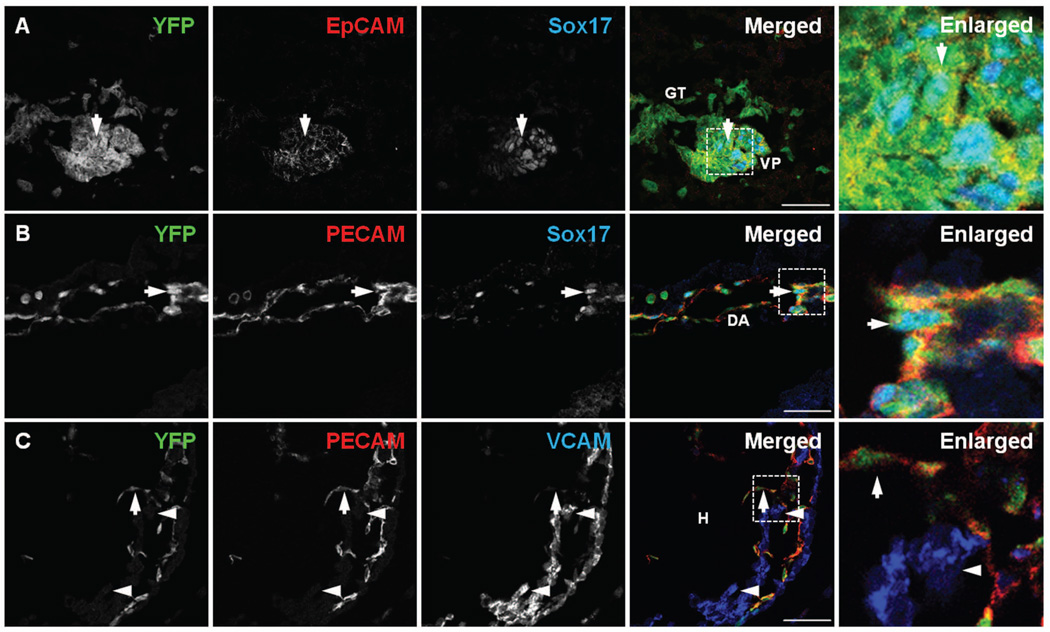
References
-
- Foster JW, Dominguez-Steglich MA, Guioli S, et al. Campomelic dysplasia and autosomal sex reversal caused by mutations in an SRY-related gene. Nature. 1994;372:525–530. - PubMed
-
- Kamachi Y, Uchikawa M, Collignon J, et al. Involvement of Sox1, 2 and 3 in the early and subsequent molecular events of lens induction. Development. 1998;125:2521–2532. - PubMed
-
- Schilham MW, Oosterwegel MA, Moerer P, et al. Defects in cardiac outflow tract formation and pro-B-lymphocyte expansion in mice lacking Sox-4. Nature. 1996;380:711–714. - PubMed
-
- Pingault V, Bondurand N, Kuhlbrodt K, et al. SOX10 mutations in patients with Waardenburg-Hirschsprung disease. Nat Genet. 1998;18:171–173. - PubMed
-
- Kanai-Azuma M, Kanai Y, Gad JM, et al. Depletion of definitive gut endoderm in Sox17-null mutant mice. Development. 2002;129:2367–2379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U19 DK072495/DK/NIDDK NIH HHS/United States
- P60 DK020593/DK/NIDDK NIH HHS/United States
- DK58404/DK/NIDDK NIH HHS/United States
- U01 DK089523/DK/NIDDK NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- U01 DK072473/DK/NIDDK NIH HHS/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- CA68485/CA/NCI NIH HHS/United States
- DK20593/DK/NIDDK NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- DK72473/DK/NIDDK NIH HHS/United States
- DK072495/DK/NIDDK NIH HHS/United States
- DK89523/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
