

The calcium/calcineurin pathway promotes hemidesmosome stability through inhibition of β4 integrin phosphorylation
- PMID: 22865863
- PMCID: PMC3463366
- DOI: 10.1074/jbc.M112.385245
The calcium/calcineurin pathway promotes hemidesmosome stability through inhibition of β4 integrin phosphorylation
Abstract
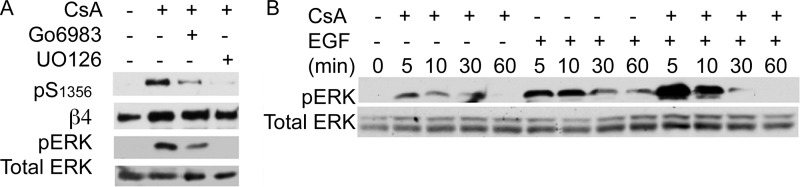
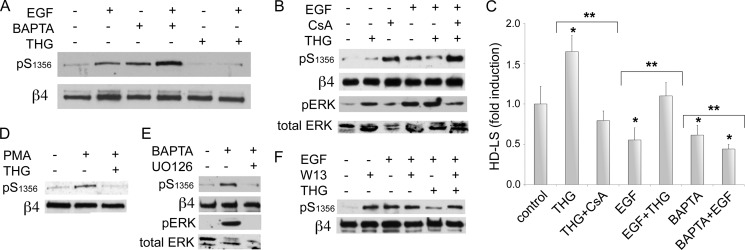
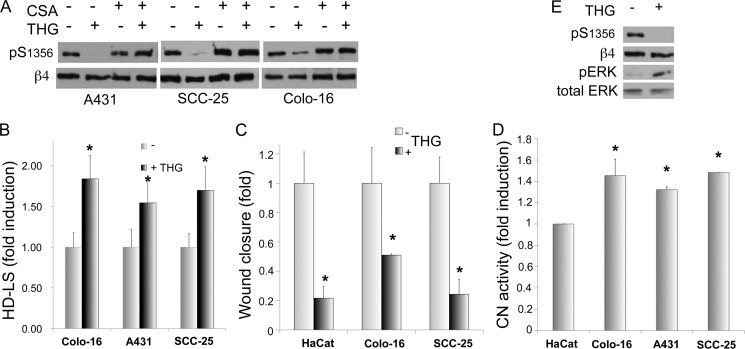
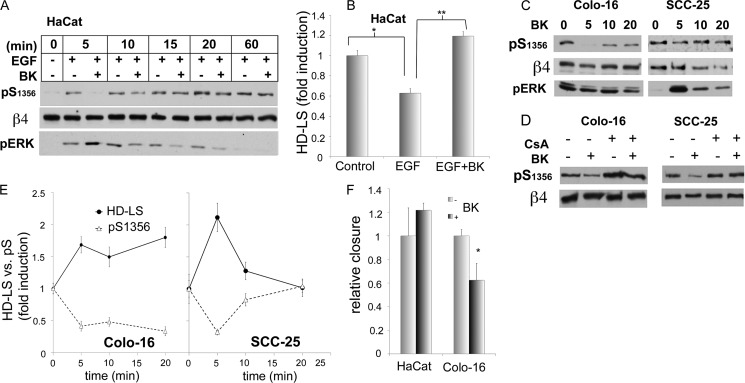
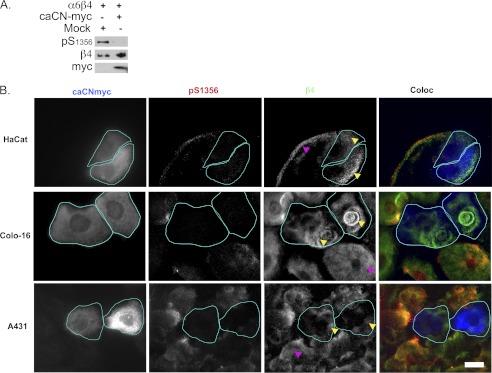
Cell migration depends on cells being able to create and disassemble adhesive contacts. Hemidesmosomes are multiprotein structures that attach epithelia to basal lamina and disassemble during migration and carcinoma invasion. Phosphorylation of the β4 integrin, a hemidesmosome component, induces disassembly. Although kinases involved in β4 phosphorylation have been identified, little is known about phosphatases countering kinase action. Here we report that calcineurin, a serine-threonine protein phosphatase, regulates β4 phosphorylation. Calcineurin inhibitor cyclosporin A (CsA) and calcineurin-siRNA increase β4 phosphorylation, induce hemidesmosome disassembly, and increase migration in HaCat keratinocytes, suggesting that calcineurin negatively regulates β4 phosphorylation. We found no direct dephosphorylation of β4 by calcineurin or association between β4 and calcineurin, suggesting indirect regulation of β4 phosphorylation. We therefore assessed calcineurin influence on MAPK and PKC, known to phosphorylate β4. CsA increased MAPK activity, whereas MAPK inhibitors reduced CsA-induced β4 phosphorylation, suggesting that calcineurin restricts β4 phosphorylation by MAPK. Calcineurin is activated by calcium. Increased [Ca(2+)](i) reduces β4 phosphorylation and stabilizes hemidesmosomes, effects that are reversed by CsA, indicating that calcineurin mediates calcium effects on β4. However, MAPK activation is increased when [Ca(2+)](i) is increased, suggesting that calcineurin activates an additional mechanism that counteracts MAPK-induced β4 phosphorylation. Interestingly, in some squamous cell carcinoma cells, which have reduced hemidesmosomes and increased β4 phosphorylation, an increase in [Ca(2+)](i) using thapsigargin, bradykinin, or acetylcholine can increase hemidesmosomes and reduce β4 phosphorylation in a calcineurin-dependent manner. These findings have implications in calcineurin-inhibitor induced carcinoma, a complication of immunosuppressive therapy.
Figures

Similar articles
-
Role of β4 integrin phosphorylation in human invasive squamous cell carcinoma: regulation of hemidesmosome stability modulates cell migration.Lab Invest. 2011 Oct;91(10):1414-26. doi: 10.1038/labinvest.2011.104. Epub 2011 Jul 18. Lab Invest. 2011. PMID: 21769085 Free PMC article.
-
Phosphorylation of threonine 1736 in the C-terminal tail of integrin β4 contributes to hemidesmosome disassembly.Mol Biol Cell. 2012 Apr;23(8):1475-85. doi: 10.1091/mbc.E11-11-0957. Epub 2012 Feb 22. Mol Biol Cell. 2012. PMID: 22357621 Free PMC article.
-
Protein kinase C-alpha phosphorylation of specific serines in the connecting segment of the beta 4 integrin regulates the dynamics of type II hemidesmosomes.Mol Cell Biol. 2004 May;24(10):4351-60. doi: 10.1128/MCB.24.10.4351-4360.2004. Mol Cell Biol. 2004. PMID: 15121854 Free PMC article.
-
Regulation of hemidesmosome disassembly by growth factor receptors.Curr Opin Cell Biol. 2008 Oct;20(5):589-96. doi: 10.1016/j.ceb.2008.05.001. Epub 2008 Jun 24. Curr Opin Cell Biol. 2008. PMID: 18583123 Review.
-
Integrin beta4, keratinocytes and papillomavirus infection.Int J Mol Med. 2006 Feb;17(2):195-202. Int J Mol Med. 2006. PMID: 16391815 Review.
Cited by
-
Molecular architecture and function of the hemidesmosome.Cell Tissue Res. 2015 May;360(2):363-78. doi: 10.1007/s00441-014-2061-z. Epub 2014 Dec 9. Cell Tissue Res. 2015. PMID: 25487405 Free PMC article. Review.
-
PKD2 and RSK1 Regulate Integrin β4 Phosphorylation at Threonine 1736.PLoS One. 2015 Nov 18;10(11):e0143357. doi: 10.1371/journal.pone.0143357. eCollection 2015. PLoS One. 2015. PMID: 26580203 Free PMC article.
-
Molecular architecture and function of the hemidesmosome.Cell Tissue Res. 2015 Jun;360(3):529-44. doi: 10.1007/s00441-015-2216-6. Epub 2015 May 29. Cell Tissue Res. 2015. PMID: 26017636 Free PMC article. Review.
-
Engineered basement membrane mimetic hydrogels to study mammary epithelial morphogenesis and invasion.bioRxiv [Preprint]. 2025 Mar 3:2025.02.28.640825. doi: 10.1101/2025.02.28.640825. bioRxiv. 2025. PMID: 40093166 Free PMC article. Preprint.
-
Cyclosporine in patients with atopic dermatitis modulates activated inflammatory pathways and reverses epidermal pathology.J Allergy Clin Immunol. 2014 Jun;133(6):1626-34. doi: 10.1016/j.jaci.2014.03.003. Epub 2014 Apr 29. J Allergy Clin Immunol. 2014. PMID: 24786238 Free PMC article.
References
-
- Borradori L., Sonnenberg A. (1996) Hemidesmosomes: roles in adhesion, signaling, and human diseases. Curr. Opin. Cell Biol. 8, 647–656 - PubMed
-
- Green K. J., Jones J. C. (1996) Desmosomes and hemidesmosomes: structure and function of molecular components. FASEB J. 10, 871–881 - PubMed
-
- Margadant C., Frijns E., Wilhelmsen K., Sonnenberg A. (2008) Regulation of hemidesmosome disassembly by growth factor receptors. Curr. Opin. Cell Biol. 20, 589–596 - PubMed
-
- Gipson I. K., Spurr-Michaud S., Tisdale A., Elwell J., Stepp M. A. (1993) Redistribution of the hemidesmosome components α6β4 integrin and bullous pemphigoid antigens during epithelial wound healing. Exp. Cell Res. 207, 86–98 - PubMed
-
- Herold-Mende C., Kartenbeck J., Tomakidi P., Bosch F. X. (2001) Metastatic growth of squamous cell carcinomas is correlated with up-regulation and redistribution of hemidesmosomal components. Cell Tissue Res. 306, 399–408 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
