

Review
doi: 10.1101/cshperspect.a012294.
mRNA localization and translational control in Drosophila oogenesis
Affiliations
- PMID: 22865893
- PMCID: PMC3475173
- DOI: 10.1101/cshperspect.a012294
Item in Clipboard
Review
mRNA localization and translational control in Drosophila oogenesis
Cold Spring Harb Perspect Biol.
.
Abstract
Localization of an mRNA species to a particular subcellular region can complement translational control mechanisms to produce a restricted spatial distribution of the protein it encodes. mRNA localization has been studied most in asymmetric cells such as budding yeast, early embryos, and neurons, but the process is likely to be more widespread. This article reviews the current state of knowledge about the mechanisms of mRNA localization and its functions in early embryonic development, focusing on Drosophila where the relevant knowledge is most advanced. Links between mRNA localization and translational control mechanisms also are examined.
Figures
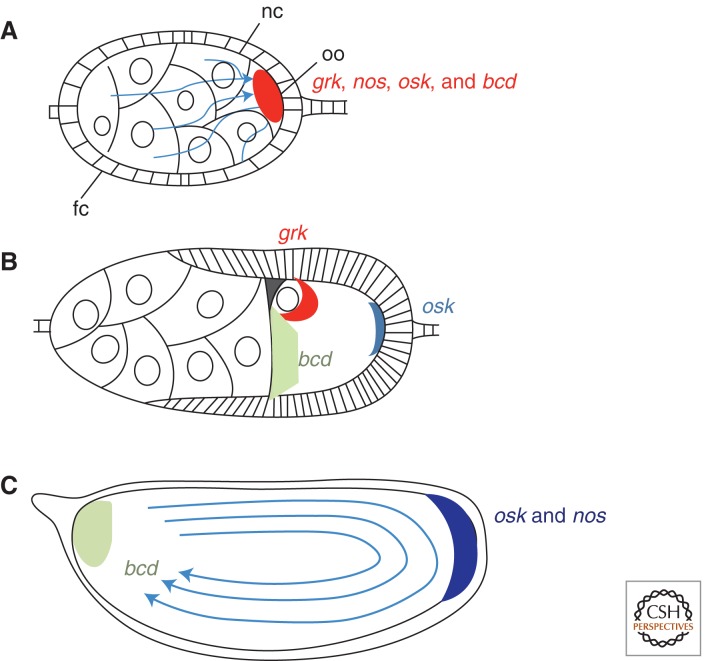
Localization of patterning mRNAs in Drosophila oogenesis. (A) In early oogenesis, several mRNAs, including grk, nos, osk, and bcd, are transported from the nurse cells through cytoplasmic bridges called ring canals into the oocyte. This involves minus-end directed transport along microtubules (blue arrows) mediated by the dynein motor complex. Abbreviations: nc, nurse cells, fc, follicle cells, oo, oocyte. (B) In mid-oogenesis, osk mRNA localizes to the posterior of the oocyte, grk mRNA localizes to the anterodorsal corner in close association with the oocyte nucleus, and bcd mRNA localizes to the anterior pole. (C) In late oogenesis, centrifugal cytoplasmic streaming (delineated by arrows) coupled with posterior anchoring brings about a further posterior enrichment of osk mRNA as well as posterior enrichment of nos mRNA. The distribution of bcd mRNA at the anterior pole is further refined.
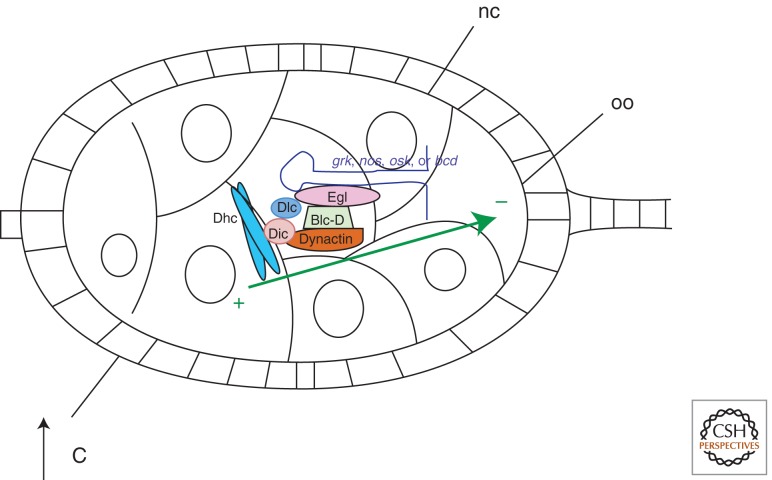
Model for linking mRNAs to the microtubule cytoskeleton for minus-end directed transport. Egalitarian (Egl) interacts directly with localization signals on mRNAs, with the carboxy-terminal end of Bicaudal-D (Bic-D), and with dynein light chain (Dlc). Bic-D interacts directly with dynactin, which in turn binds to dynein through its intermediate chain (Dic). Dynein heavy chain (Dhc) interacts with microtubules (green arrow) and catalyzes movement toward the minus-end. Although both in vivo and in vitro evidence exists to support this model for some instances of dynein-directed minus-end transport, and Egl and Bic-D are required for accumulation of grk, nos, osk, and bcd mRNAs into the oocyte, it has not yet been directly shown that this mechanism governs this particular localization event.
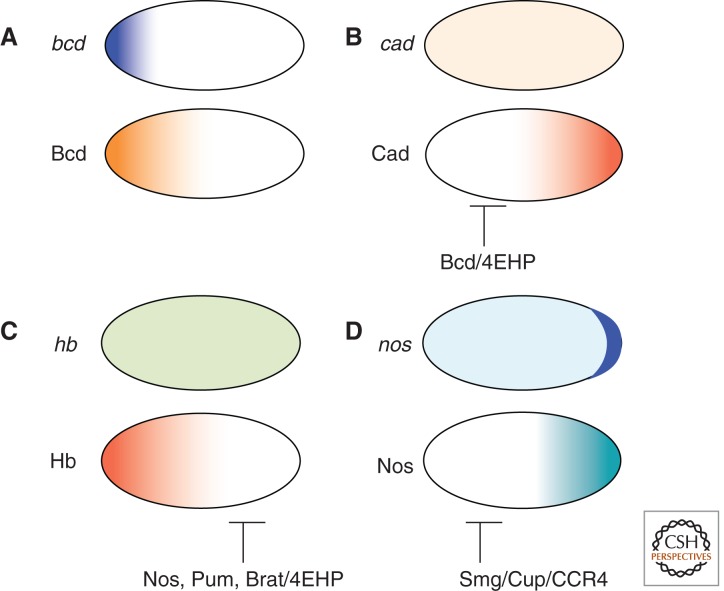
Mechanisms of establishing protein gradients in the early embryo prior to the onset of zygotic transcription. (A) Maternally expressed bcd mRNA (top panel) is localized in a steep gradient at the anterior pole. Bcd protein (lower panel) is translated from that localized mRNA and diffuses toward the posterior. (B) Maternally-expressed cad mRNA (top panel) is uniformly distributed. Translation of cad mRNA is, however, repressed by Bcd-mediated recruitment of 4EHP, resulting in a posterior-to-anterior gradient of Cad protein (lower panel) that is a mirror image of the Bcd gradient. (C) Maternally expressed hb mRNA (top panel) is uniformly distributed. Translation of hb mRNA is repressed by a complex of Nos, Pum, and Brat, that recruits 4EHP and probably other negative regulators to restrict Hb protein (lower panel) to the anterior half of the embryo. (D) Maternally expressed nos mRNA (top panel) is enriched at the posterior pole but present elsewhere. Translation of nos outside the posterior is repressed by Smg, which can recruit the repressor protein Cup and also the CCR4 deadenylase complex. Unlocalized nos is also targeted by piRNAs (not shown). Nos protein (lower panel) is translated from posteriorly-localized nos that is protected from degradation and repression.
Similar articles
-
Localization, anchoring and translational control of oskar, gurken, bicoid and nanos mRNA during Drosophila oogenesis.Fly (Austin). 2009 Jan-Mar;3(1):15-28. doi: 10.4161/fly.3.1.7751. Epub 2009 Jan 2. Fly (Austin). 2009. PMID: 19182536 Review.
-
Translational regulation and RNA localization in Drosophila oocytes and embryos.Annu Rev Genet. 2001;35:365-406. doi: 10.1146/annurev.genet.35.102401.090756. Annu Rev Genet. 2001. PMID: 11700288 Review.
-
Setting the stage for development: mRNA translation and stability during oocyte maturation and egg activation in Drosophila.Dev Dyn. 2005 Mar;232(3):593-608. doi: 10.1002/dvdy.20297. Dev Dyn. 2005. PMID: 15704150 Review.
-
Drosophila Ik2, a member of the I kappa B kinase family, is required for mRNA localization during oogenesis.Development. 2006 Apr;133(8):1467-75. doi: 10.1242/dev.02318. Epub 2006 Mar 15. Development. 2006. PMID: 16540511
-
Role of Adducin-like (hu-li tai shao) mRNA and protein localization in regulating cytoskeletal structure and function during Drosophila Oogenesis and early embryogenesis.Dev Genet. 1996;19(3):249-57. doi: 10.1002/(SICI)1520-6408(1996)19:3<249::AID-DVG8>3.0.CO;2-9. Dev Genet. 1996. PMID: 8952067
Cited by
-
Optimal RNA binding by Egalitarian, a Dynein cargo adaptor, is critical for maintaining oocyte fate in Drosophila.RNA Biol. 2021 Dec;18(12):2376-2389. doi: 10.1080/15476286.2021.1914422. Epub 2021 Apr 27. RNA Biol. 2021. PMID: 33904382 Free PMC article.
-
Gatekeeper function for Short stop at the ring canals of the Drosophila ovary.Curr Biol. 2021 Aug 9;31(15):3207-3220.e4. doi: 10.1016/j.cub.2021.05.010. Epub 2021 Jun 4. Curr Biol. 2021. PMID: 34089646 Free PMC article.
-
Translational activation of oskar mRNA: reevaluation of the role and importance of a 5' regulatory element [corrected].PLoS One. 2015 May 4;10(5):e0125849. doi: 10.1371/journal.pone.0125849. eCollection 2015. PLoS One. 2015. PMID: 25938537 Free PMC article.
-
Distinct biogenesis pathways may have led to functional divergence of the human and Drosophila Arglu1 sisRNA.EMBO Rep. 2023 Feb 6;24(2):e54350. doi: 10.15252/embr.202154350. Epub 2022 Dec 19. EMBO Rep. 2023. PMID: 36533631 Free PMC article.
-
Asymmetric distribution of biomolecules of maternal origin in the Xenopus laevis egg and their impact on the developmental plan.Sci Rep. 2018 May 29;8(1):8315. doi: 10.1038/s41598-018-26592-1. Sci Rep. 2018. PMID: 29844480 Free PMC article.
References
-
- Arnott S, Hukins DW, Dover SD 1972. Optimised parameters for RNA double-helices. Biochem Biophys Res Commun 48: 1392–1399 - PubMed
-
- Bashirullah A, Halsell SR, Cooperstock RL, Kloc M, Karaiskakis A, Fisher WW, Fu W, Hamilton JK, Etkin LD, Lipshitz HD 1999. Joint action of two RNA degradation pathways controls the timing of maternal transcript elimination at the midblastula transition in Drosophila melanogaster. EMBO J 18: 2610–2620 - PMC - PubMed
-
- Bastock R, St Johnston D 2008. Drosophila oogenesis. Curr Biol 18: R1082–R1087 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases