

The evolving concept of cancer and metastasis stem cells
- PMID: 22869594
- PMCID: PMC3413352
- DOI: 10.1083/jcb.201202014
The evolving concept of cancer and metastasis stem cells
Abstract
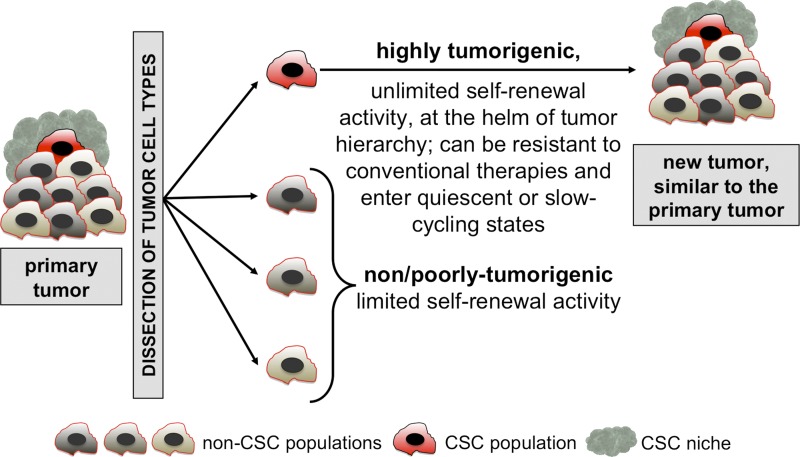
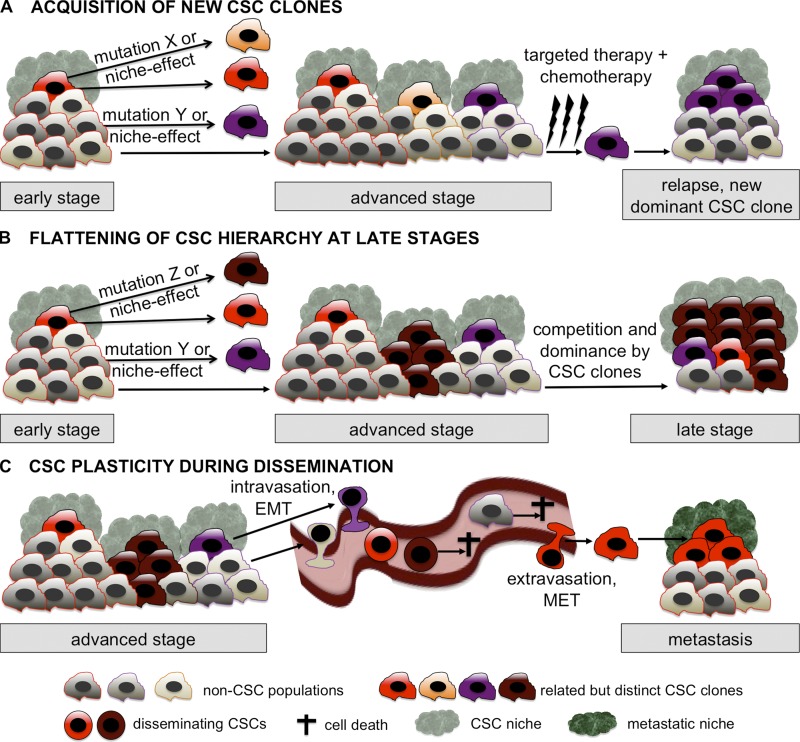
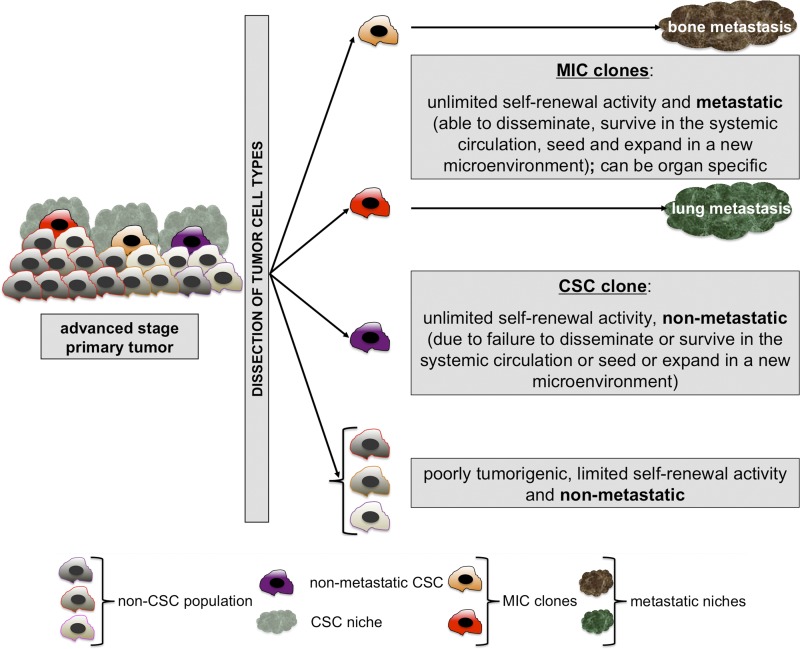
The cancer stem cell (CSC) concept, which arose more than a decade ago, proposed that tumor growth is sustained by a subpopulation of highly malignant cancerous cells. These cells, termed CSCs, comprise the top of the tumor cell hierarchy and have been isolated from many leukemias and solid tumors. Recent work has discovered that this hierarchy is embedded within a genetically heterogeneous tumor, in which various related but distinct subclones compete within the tumor mass. Thus, genetically distinct CSCs exist on top of each subclone, revealing a highly complex cellular composition of tumors. The CSC concept has therefore evolved to better model the complex and highly dynamic processes of tumorigenesis, tumor relapse, and metastasis.
Figures
References
-
- American Cancer Society 2011. Cancer Facts & Figures 2011. American Cancer Society, Atlanta: 58 pp
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
