

Spatial ordering of chromosomes enhances the fidelity of chromosome partitioning in cyanobacteria
- PMID: 22869746
- PMCID: PMC3427121
- DOI: 10.1073/pnas.1211144109
Spatial ordering of chromosomes enhances the fidelity of chromosome partitioning in cyanobacteria
Abstract
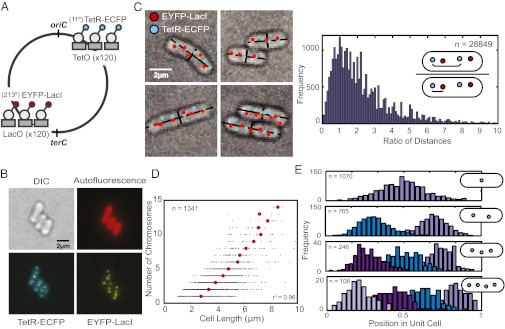
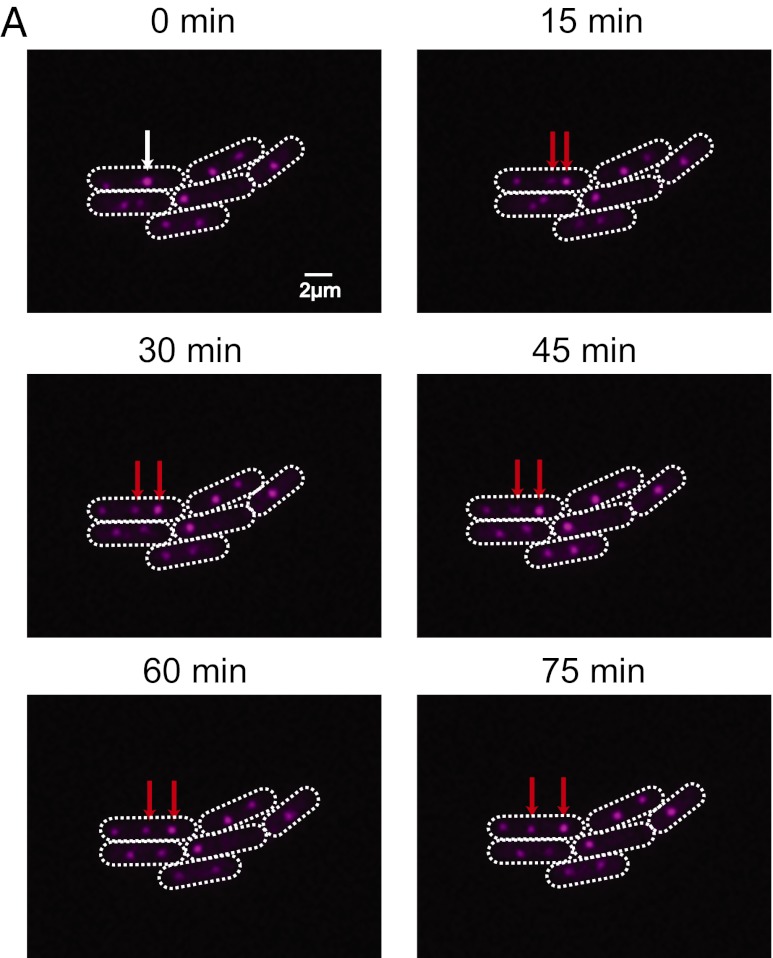
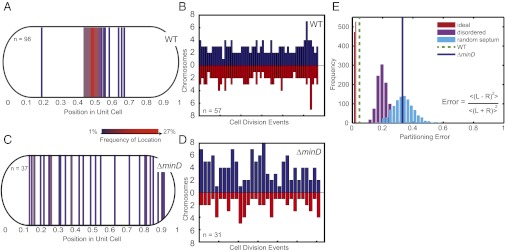
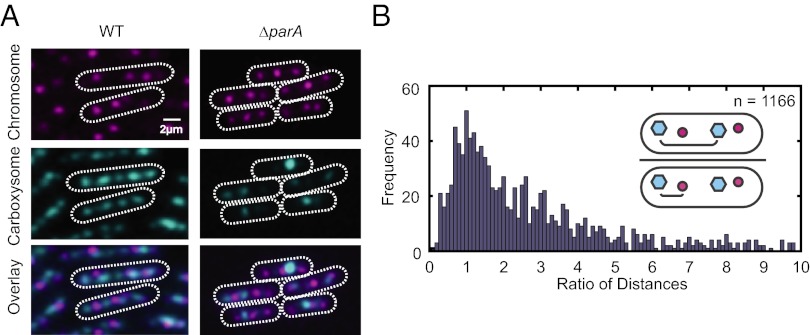
Many cyanobacteria have been shown to harbor multiple chromosome copies per cell, yet little is known about the organization, replication, and segregation of these chromosomes. Here, we visualize individual chromosomes in the cyanobacterium Synechococcus elongatus via time-lapse fluorescence microscopy. We find that chromosomes are equally spaced along the long axis of the cell and are interspersed with another regularly spaced subcellular compartment, the carboxysome. This remarkable organization of the cytoplasm along with accurate midcell septum placement allows for near-optimal segregation of chromosomes to daughter cells. Disruption of either chromosome ordering or midcell septum placement significantly increases the chromosome partitioning error. We find that chromosome replication is both asynchronous and independent of the position of the chromosome in the cell and that spatial organization is preserved after replication. Our findings on chromosome organization, replication, and segregation in S. elongatus provide a basis for understanding chromosome dynamics in bacteria with multiple chromosomes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
