

L-citrulline supplementation reverses the impaired airway relaxation in neonatal rats exposed to hyperoxia
- PMID: 22870905
- PMCID: PMC3487946
- DOI: 10.1186/1465-9921-13-68
L-citrulline supplementation reverses the impaired airway relaxation in neonatal rats exposed to hyperoxia
Abstract
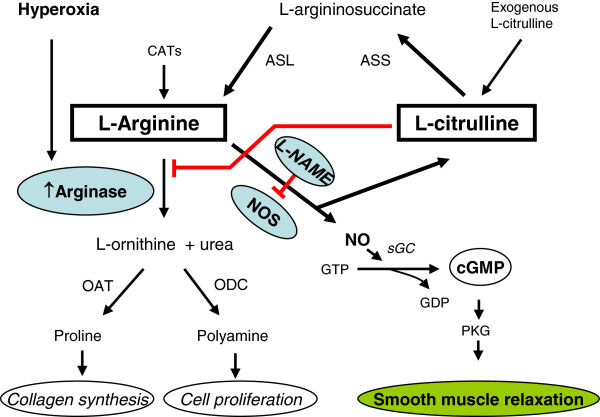
Background: Hyperoxia is shown to impair airway relaxation via limiting L-arginine bioavailability to nitric oxide synthase (NOS) and reducing NO production as a consequence. L-arginine can also be synthesized by L-citrulline recycling. The role of L-citrulline supplementation was investigated in the reversing of hyperoxia-induced impaired relaxation of rat tracheal smooth muscle (TSM).
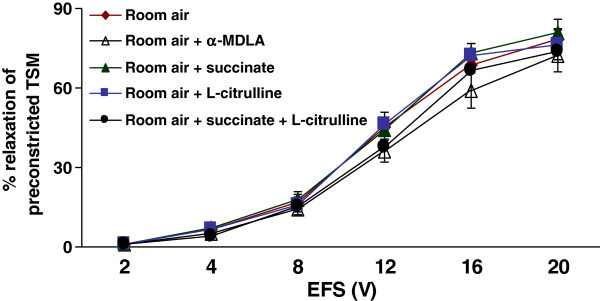
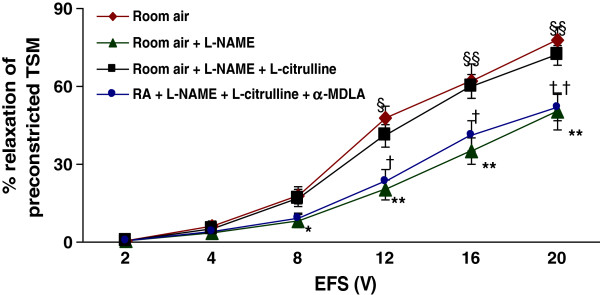
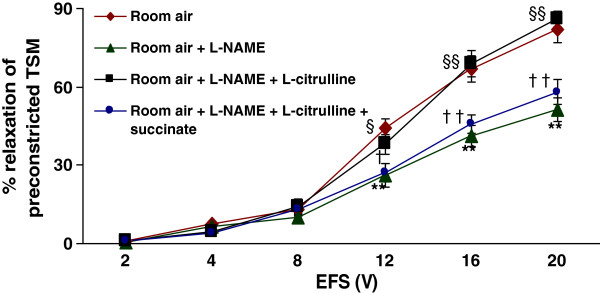
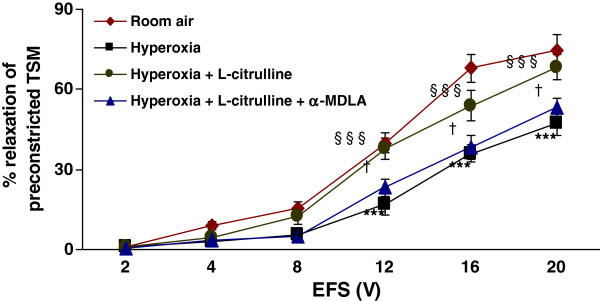
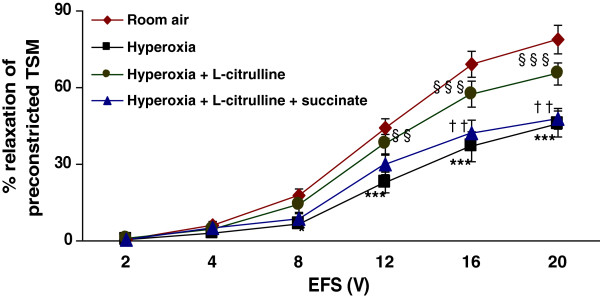
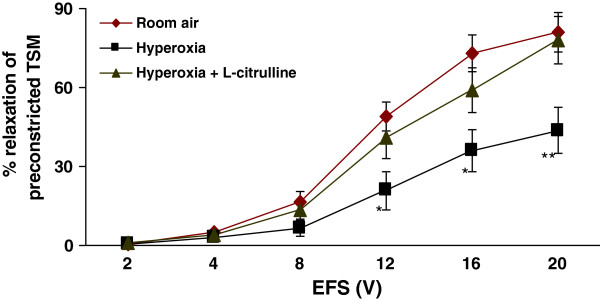
Methods: Electrical field stimulation (EFS, 2-20 V)-induced relaxation was measured under in vitro conditions in preconstricted tracheal preparations obtained from 12 day old rat pups exposed to room air or hyperoxia (>95% oxygen) for 7 days supplemented with L-citrulline or saline (in vitro or in vivo). The role of the L-citrulline/L-arginine cycle under basal conditions was studied by incubation of preparations in the presence of argininosuccinate synthase (ASS) inhibitor [α-methyl-D, L-aspartate, 1 mM] or argininosuccinate lyase inhibitor (ASL) succinate (1 mM) and/or NOS inhibitor [Nω-nitro-L-arginine methyl ester; 100 μM] with respect to the presence or absence of L-citrulline (2 mM).
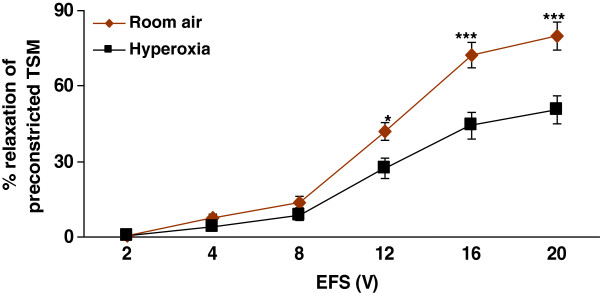
Results: Hyperoxia impaired the EFS-induced relaxation of TSM as compared to room air control (p < 0.001; 0.5 ± 0.1% at 2 V to 50.6 ± 5.7% at 20 V in hyperoxic group: 0.7 ± 0.2 at 2 V to 80.0 ± 5.6% at 20 V in room air group). Inhibition of ASS or ASL, and L-citrulline supplementation did not affect relaxation responses under basal conditions. However, inhibition of NOS significantly reduced relaxation responses (p < 0.001), which were restored to control level by L-citrulline. L-citrulline supplementation in vivo and in vitro also reversed the hyperoxia-impaired relaxation. The differences were significant (p <0.001; 0.8 ± 0.3% at 2 V to 47.1 ± 4.1% at 20 V without L-citrulline; 0.9 ± 0.3% at 2 V to 68.2 ± 4.8% at 20 V with L-citrulline). Inhibition of ASS or ASL prevented this effect of L-citrulline.
Conclusion: The results indicate the presence of an L-citrulline/L-arginine cycle in the airways of rat pups. L-citrulline recycling does not play a major role under basal conditions in airways, but it has an important role under conditions of substrate limitations to NOS as a source of L-arginine, and L-citrulline supplementation reverses the impaired relaxation of airways under hyperoxic conditions.
Figures
Similar articles
-
Role of the L-citrulline/L-arginine cycle in iNANC nerve-mediated nitric oxide production and airway smooth muscle relaxation in allergic asthma.Eur J Pharmacol. 2006 Sep 28;546(1-3):171-6. doi: 10.1016/j.ejphar.2006.07.041. Epub 2006 Jul 27. Eur J Pharmacol. 2006. PMID: 16919264
-
Role of arginase in impairing relaxation of lung parenchyma of hyperoxia-exposed neonatal rats.Neonatology. 2012;101(2):106-15. doi: 10.1159/000329540. Epub 2011 Sep 23. Neonatology. 2012. PMID: 21952491 Free PMC article.
-
Curcumin analogs (B2BrBC and C66) supplementation attenuates airway hyperreactivity and promote airway relaxation in neonatal rats exposed to hyperoxia.Physiol Rep. 2020 Aug;8(16):e14555. doi: 10.14814/phy2.14555. Physiol Rep. 2020. PMID: 32812392 Free PMC article.
-
Supplementation with Nitric Oxide Precursors for Strength Performance: A Review of the Current Literature.Nutrients. 2023 Jan 28;15(3):660. doi: 10.3390/nu15030660. Nutrients. 2023. PMID: 36771366 Free PMC article. Review.
-
[The effect of supplementation with L-arginine and L-citrulline on physical performance: a systematic review].Nutr Hosp. 2019 Dec 26;36(6):1389-1402. doi: 10.20960/nh.02478. Nutr Hosp. 2019. PMID: 31682459 Spanish.
Cited by
-
VIP/PACAP signaling as an alternative target during hyperoxic exposure in preterm newborns.Physiol Res. 2021 Aug 31;70(4):489-499. doi: 10.33549/physiolres.934638. Epub 2021 Jun 1. Physiol Res. 2021. PMID: 34062074 Free PMC article. Review.
-
Targeting the lung endothelial niche to promote angiogenesis and regeneration: A review of applications.Front Mol Biosci. 2022 Dec 19;9:1093369. doi: 10.3389/fmolb.2022.1093369. eCollection 2022. Front Mol Biosci. 2022. PMID: 36601582 Free PMC article. Review.
-
The Effect of L-Citrulline Supplementation on Outcomes of Critically Ill Patients under Mechanical Ventilation; a Double-Blind Randomized Controlled Trial.Arch Acad Emerg Med. 2023 Jan 1;11(1):e11. doi: 10.22037/aaem.v11i1.1774. eCollection 2023. Arch Acad Emerg Med. 2023. PMID: 36620736 Free PMC article.
-
Protective Effects of Resveratrol Against Airway Hyperreactivity, Oxidative Stress, and Lung Inflammation in a Rat Pup Model of Bronchopulmonary Dysplasia.Physiol Res. 2024 Apr 30;73(2):239-251. doi: 10.33549/physiolres.935239. Physiol Res. 2024. PMID: 38710061 Free PMC article.
-
Arginine Therapy for Lung Diseases.Front Pharmacol. 2021 Mar 23;12:627503. doi: 10.3389/fphar.2021.627503. eCollection 2021. Front Pharmacol. 2021. PMID: 33833679 Free PMC article. Review.
References
-
- Pelkonen AS, Hakulinen AL, Turpeinen M. Bronchial lability and responsiveness in school children born very preterm. Am J Respir Crit Care Med. 1997;156:1178–1184. - PubMed
-
- Hack M, Taylor HG, Drotar D, Schluchter M, Cartar L, Andreias L, Wilson-Costello D, Klein N. Chronic conditions, functional limitations and special health care needs of school-aged children born with extremely low-birth-weight in the 1990's. JAMA. 2005;294:318–325. doi: 10.1001/jama.294.3.318. - DOI - PubMed
-
- Sopi RB, Martin RJ, Haxhiu MA, Dreshaj IA, Yao Q, Jafri A, Zaidi SI. Role of brain-derived neurotrophic factor in hyperoxia-induced enhancement of contractility and impairment of relaxation in lung parenchyma. Am J Physiol Lung Cell Mol Physiol. 2008;295:L348–L355. doi: 10.1152/ajplung.00067.2008. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
