

Neonatal exposure to the cyanobacterial toxin BMAA induces changes in protein expression and neurodegeneration in adult hippocampus
- PMID: 22872059
- PMCID: PMC3498744
- DOI: 10.1093/toxsci/kfs241
Neonatal exposure to the cyanobacterial toxin BMAA induces changes in protein expression and neurodegeneration in adult hippocampus
Erratum in
- Toxicol Sci. 2013 Mar;132(1):251
Abstract
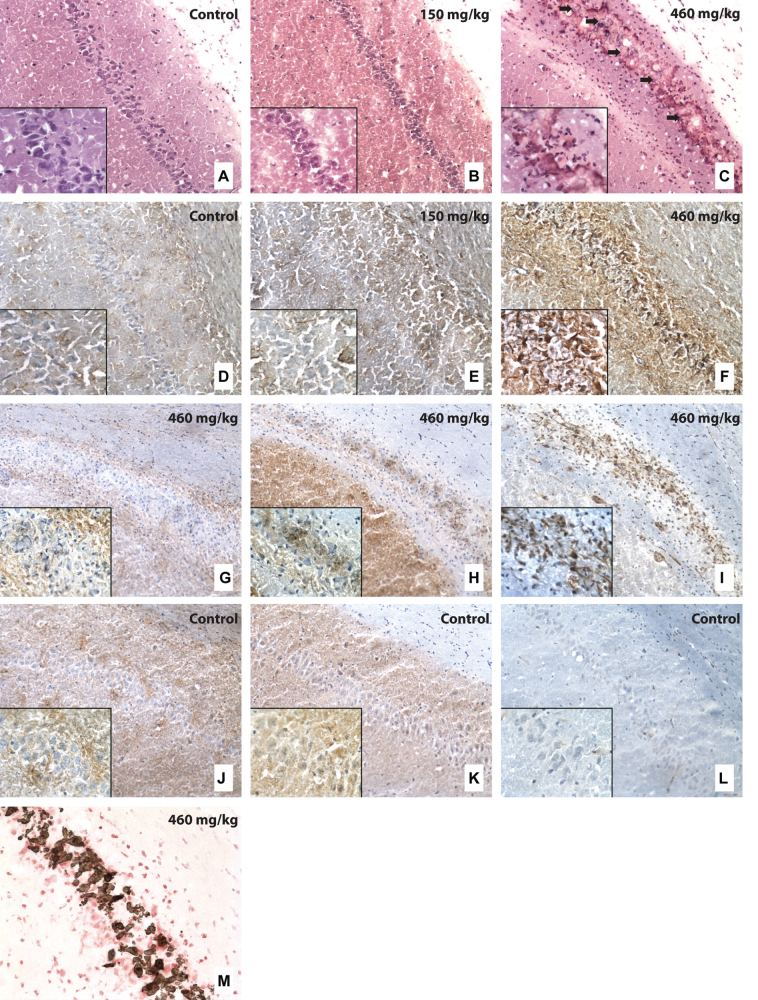
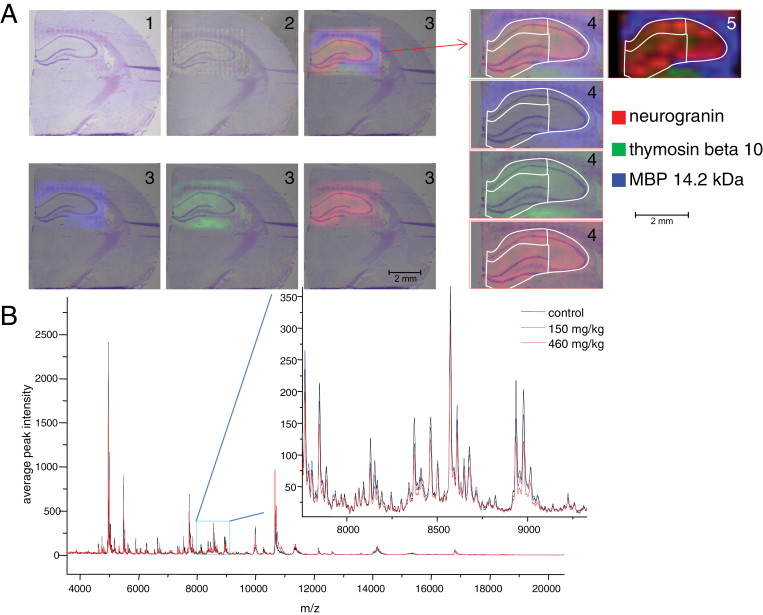
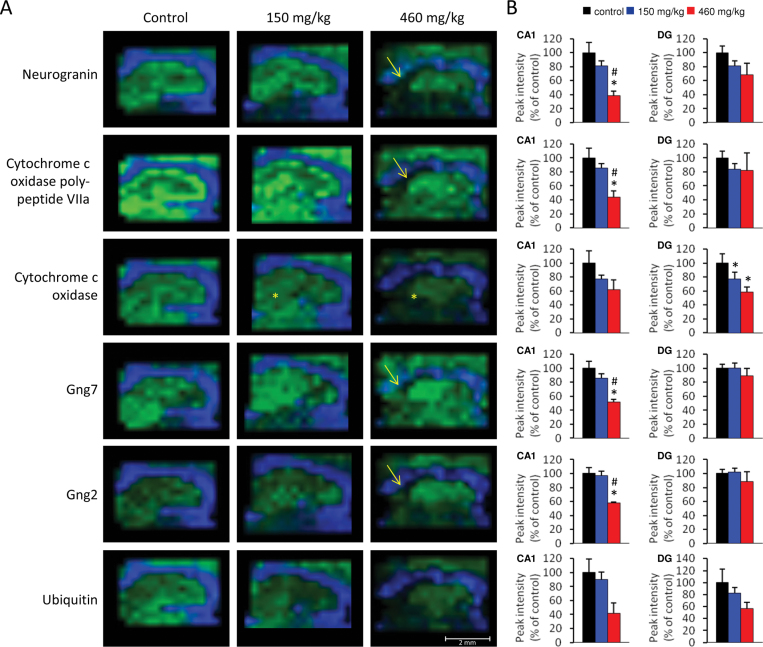
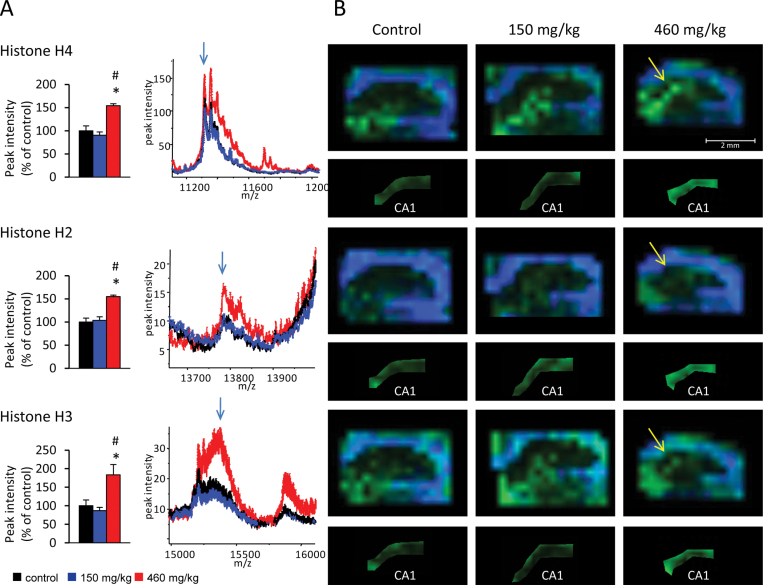
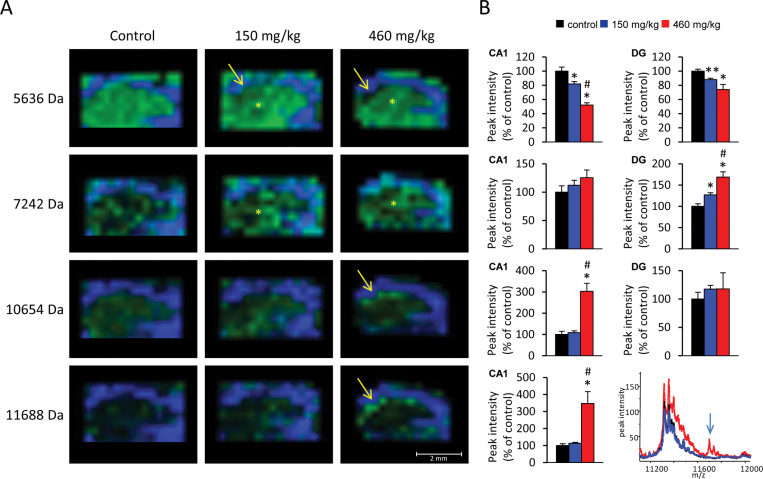
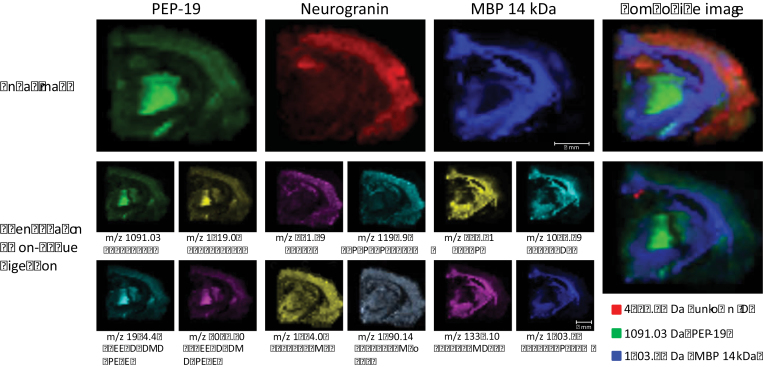
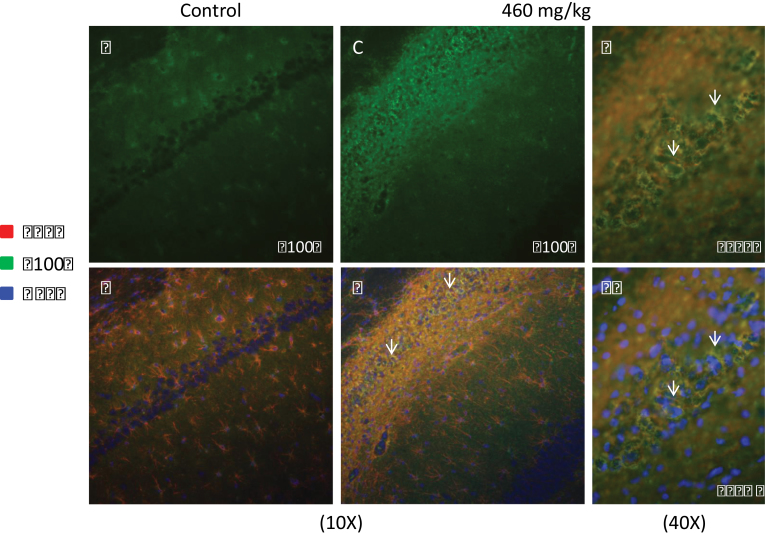
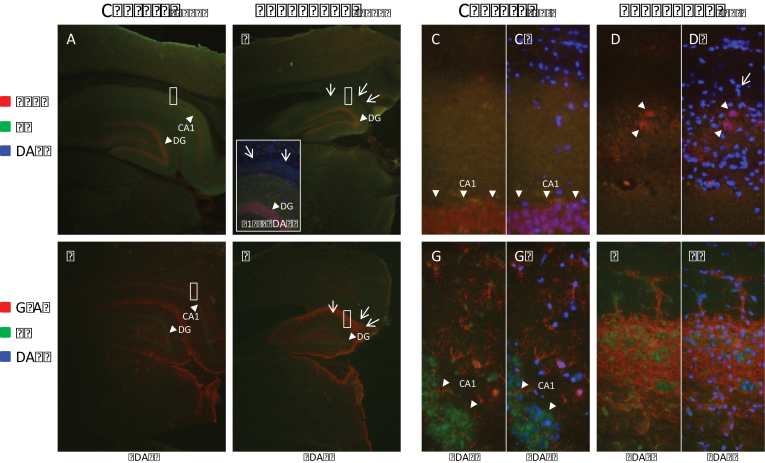
The cyanobacterial toxin β-N-methylamino-L-alanine (BMAA) has been proposed to contribute to neurodegenerative disease. We have previously reported a selective uptake of BMAA in the mouse neonatal hippocampus and that exposure during the neonatal period causes learning and memory impairments in adult rats. The aim of this study was to characterize effects in the brain of 6-month-old rats treated neonatally (postnatal days 9-10) with the glutamatergic BMAA. Protein changes were examined using the novel technique Matrix-Assisted Laser Desorption Ionization (MALDI) imaging mass spectrometry (IMS) for direct imaging of proteins in brain cryosections, and histological changes were examined using immunohistochemistry and histopathology. The results showed long-term changes including a decreased expression of proteins involved in energy metabolism and intracellular signaling in the adult hippocampus at a dose (150 mg/kg) that gave no histopathological lesions in this brain region. Developmental exposure to a higher dose (460 mg/kg) also induced changes in the expression of S100β, histones, calcium- and calmodulin-binding proteins, and guanine nucleotide-binding proteins. At this dose, severe lesions in the adult hippocampus including neuronal degeneration, cell loss, calcium deposits, and astrogliosis were evident. The data demonstrate subtle, sometimes dose-dependent, but permanent effects of a lower neonatal dose of BMAA in the adult hippocampus suggesting that BMAA could potentially disturb many processes during the development. The detection of BMAA in seafood stresses the importance of evaluating the magnitude of human exposure to this neurotoxin.
Figures
Similar articles
-
Quality measures of imaging mass spectrometry aids in revealing long-term striatal protein changes induced by neonatal exposure to the cyanobacterial toxin β-N-methylamino-L-alanine (BMAA).Mol Cell Proteomics. 2014 Jan;13(1):93-104. doi: 10.1074/mcp.M113.031435. Epub 2013 Oct 14. Mol Cell Proteomics. 2014. PMID: 24126143 Free PMC article.
-
MALDI imaging delineates hippocampal glycosphingolipid changes associated with neurotoxin induced proteopathy following neonatal BMAA exposure.Biochim Biophys Acta Proteins Proteom. 2017 Jul;1865(7):740-746. doi: 10.1016/j.bbapap.2016.12.004. Epub 2016 Dec 9. Biochim Biophys Acta Proteins Proteom. 2017. PMID: 27956354 Free PMC article.
-
Early hippocampal cell death, and late learning and memory deficits in rats exposed to the environmental toxin BMAA (β-N-methylamino-L-alanine) during the neonatal period.Behav Brain Res. 2011 Jun 1;219(2):310-20. doi: 10.1016/j.bbr.2011.01.056. Epub 2011 Feb 12. Behav Brain Res. 2011. PMID: 21315110
-
Mechanisms and Effects Posed by Neurotoxic Products of Cyanobacteria/Microbial Eukaryotes/Dinoflagellates in Algae Blooms: a Review.Neurotox Res. 2018 Jan;33(1):153-167. doi: 10.1007/s12640-017-9780-3. Epub 2017 Aug 23. Neurotox Res. 2018. PMID: 28836116 Review.
-
Mechanisms of beta-N-methylamino-L-alanine induced neurotoxicity.Amyotroph Lateral Scler. 2009;10 Suppl 2:56-60. doi: 10.3109/17482960903269062. Amyotroph Lateral Scler. 2009. PMID: 19929733 Review.
Cited by
-
Current status and future perspectives of mass spectrometry imaging.Int J Mol Sci. 2013 May 28;14(6):11277-301. doi: 10.3390/ijms140611277. Int J Mol Sci. 2013. PMID: 23759983 Free PMC article. Review.
-
Quality measures of imaging mass spectrometry aids in revealing long-term striatal protein changes induced by neonatal exposure to the cyanobacterial toxin β-N-methylamino-L-alanine (BMAA).Mol Cell Proteomics. 2014 Jan;13(1):93-104. doi: 10.1074/mcp.M113.031435. Epub 2013 Oct 14. Mol Cell Proteomics. 2014. PMID: 24126143 Free PMC article.
-
Creating a Simian Model of Guam ALS/PDC Which Reflects Chamorro Lifetime BMAA Exposures.Neurotox Res. 2018 Jan;33(1):24-32. doi: 10.1007/s12640-017-9745-6. Epub 2017 May 6. Neurotox Res. 2018. PMID: 28478528
-
Β-N-Methylamino-L-Alanine (BMAA) Toxicity Is Gender and Exposure-Age Dependent in Rats.Toxins (Basel). 2017 Dec 27;10(1):16. doi: 10.3390/toxins10010016. Toxins (Basel). 2017. PMID: 29280981 Free PMC article.
-
Correlated Materials Characterization via Multimodal Chemical and Functional Imaging.ACS Nano. 2018 Dec 26;12(12):11798-11818. doi: 10.1021/acsnano.8b07292. Epub 2018 Dec 3. ACS Nano. 2018. PMID: 30422627 Free PMC article.
References
-
- Aerni H. R., Cornett D. S., Caprioli R. M. (2006). Automated acoustic matrix deposition for MALDI sample preparation. Anal. Chem. 78 827–834 - PubMed
-
- Andersson M., Groseclose M. R., Deutch A. Y., Caprioli R. M. (2008). Imaging mass spectrometry of proteins and peptides: 3D volume reconstruction. Nat. Methods 5 101–108 - PubMed
-
- Banack S. A., Cox P. A. (2003). Biomagnification of cycad neurotoxins in flying foxes: Implications for ALS-PDC in Guam. Neurology 61 387–389 - PubMed
-
- Barger S. W., Van Eldik L. J. (1992). S100 beta stimulates calcium fluxes in glial and neuronal cells. J. Biol. Chem. 267 9689–9694 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
