

The application of information theory to biochemical signaling systems
- PMID: 22872091
- PMCID: PMC3820280
- DOI: 10.1088/1478-3975/9/4/045011
The application of information theory to biochemical signaling systems
Abstract
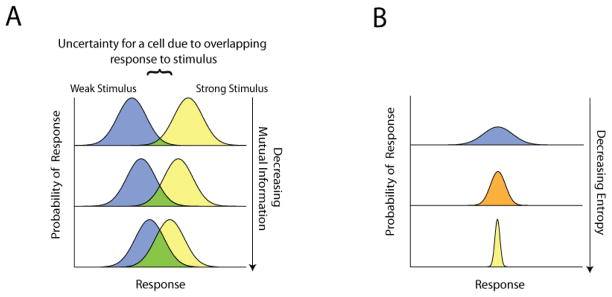
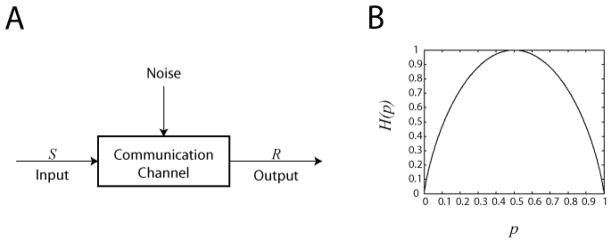
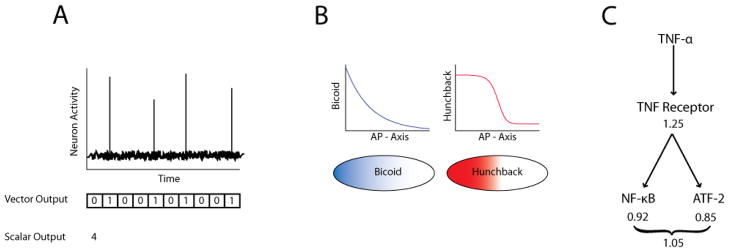
Cell signaling can be thought of fundamentally as an information transmission problem in which chemical messengers relay information about the external environment to the decision centers within a cell. Due to the biochemical nature of cellular signal transduction networks, molecular noise will inevitably limit the fidelity of any messages received and processed by a cell's signal transduction networks, leaving it with an imperfect impression of its environment. Fortunately, Shannon's information theory provides a mathematical framework independent of network complexity that can quantify the amount of information that can be transmitted despite biochemical noise. In particular, the channel capacity can be used to measure the maximum number of stimuli a cell can distinguish based upon the noisy responses of its signaling systems. Here, we provide a primer for quantitative biologists that covers fundamental concepts of information theory, highlights several key considerations when experimentally measuring channel capacity, and describes successful examples of the application of information theoretic analysis to biological signaling.
Figures
Similar articles
-
Information theory and the ethylene genetic network.Plant Signal Behav. 2011 Oct;6(10):1483-98. doi: 10.4161/psb.6.10.16424. Epub 2011 Oct 1. Plant Signal Behav. 2011. PMID: 21897127 Free PMC article. Review.
-
Information theory and signal transduction systems: from molecular information processing to network inference.Semin Cell Dev Biol. 2014 Nov;35:98-108. doi: 10.1016/j.semcdb.2014.06.011. Epub 2014 Jun 19. Semin Cell Dev Biol. 2014. PMID: 24953199 Review.
-
Analysis of cellular signal transduction from an information theoretic approach.Semin Cell Dev Biol. 2016 Mar;51:24-31. doi: 10.1016/j.semcdb.2015.12.011. Epub 2015 Dec 19. Semin Cell Dev Biol. 2016. PMID: 26710655 Review.
-
Design and analysis of molecular relay channels: an information theoretic approach.IEEE Trans Nanobioscience. 2010 Sep;9(3):213-21. doi: 10.1109/TNB.2010.2050070. Epub 2010 Jun 3. IEEE Trans Nanobioscience. 2010. PMID: 20525537
-
Information processing in bacteria: memory, computation, and statistical physics: a key issues review.Rep Prog Phys. 2016 May;79(5):052601. doi: 10.1088/0034-4885/79/5/052601. Epub 2016 Apr 8. Rep Prog Phys. 2016. PMID: 27058315 Free PMC article. Review.
Cited by
-
Tunable signal processing through a kinase control cycle: the IKK signaling node.Biophys J. 2013 Jul 2;105(1):231-41. doi: 10.1016/j.bpj.2013.05.013. Biophys J. 2013. PMID: 23823243 Free PMC article.
-
Epigenetics as a mediator of plasticity in cancer.Science. 2023 Feb 10;379(6632):eaaw3835. doi: 10.1126/science.aaw3835. Epub 2023 Feb 10. Science. 2023. PMID: 36758093 Free PMC article. Review.
-
Exploring the information transmission properties of noise-induced dynamics: application to glioma differentiation.BMC Bioinformatics. 2019 Jul 4;20(1):375. doi: 10.1186/s12859-019-2970-7. BMC Bioinformatics. 2019. PMID: 31272368 Free PMC article.
-
Systems biology: the role of engineering in the reverse engineering of biological signaling.Cells. 2013 May 31;2(2):393-413. doi: 10.3390/cells2020393. Cells. 2013. PMID: 24709707 Free PMC article.
-
Information processing in the adaptation of Saccharomyces cerevisiae to osmotic stress: an analysis of the phosphorelay system.Syst Synth Biol. 2014 Dec;8(4):297-306. doi: 10.1007/s11693-014-9146-6. Epub 2014 Apr 19. Syst Synth Biol. 2014. PMID: 26396653 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources