

Essential anaplerotic role for the energy-converting hydrogenase Eha in hydrogenotrophic methanogenesis
- PMID: 22872868
- PMCID: PMC3458328
- DOI: 10.1073/pnas.1208779109
Essential anaplerotic role for the energy-converting hydrogenase Eha in hydrogenotrophic methanogenesis
Abstract
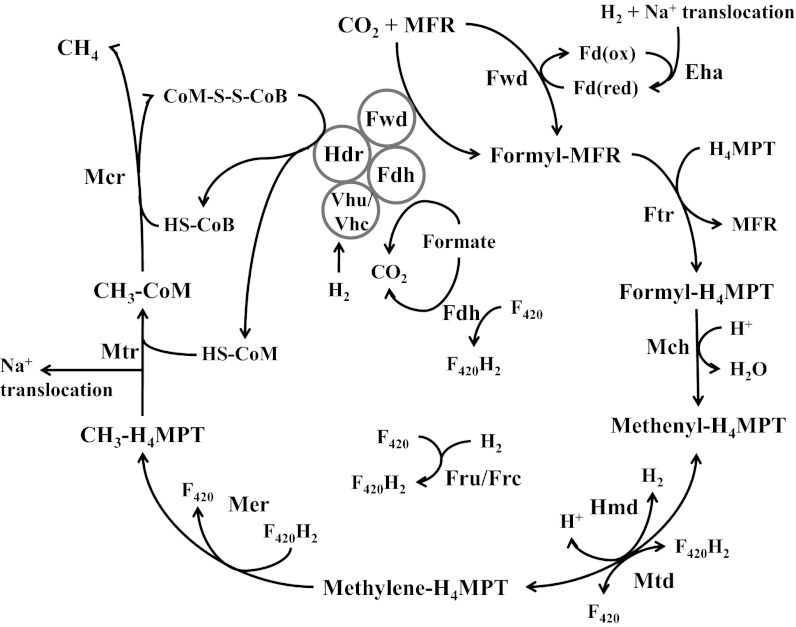
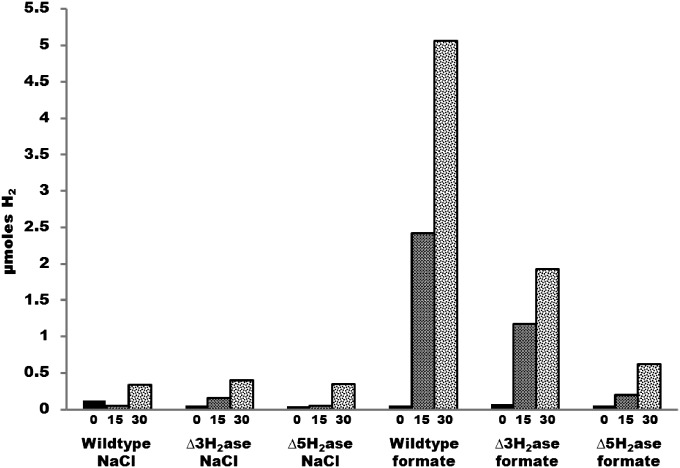
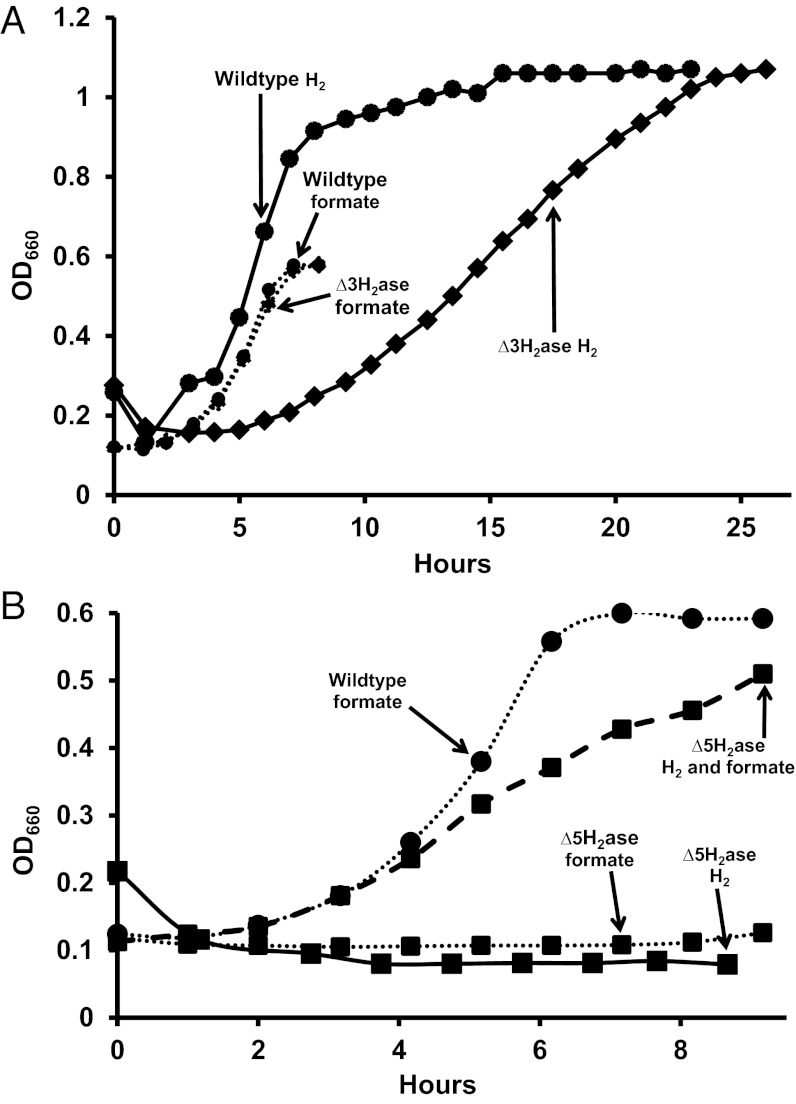
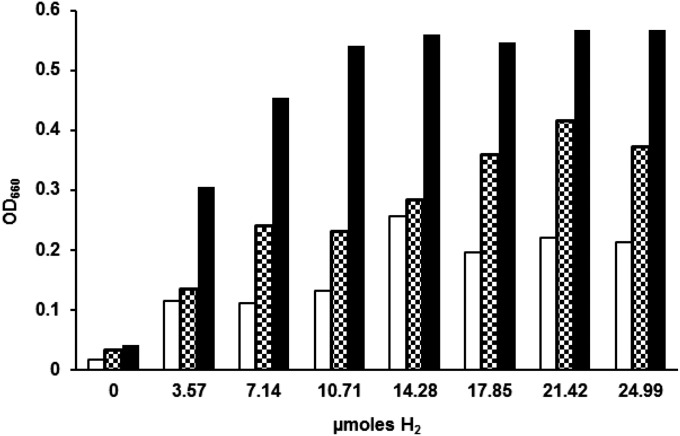
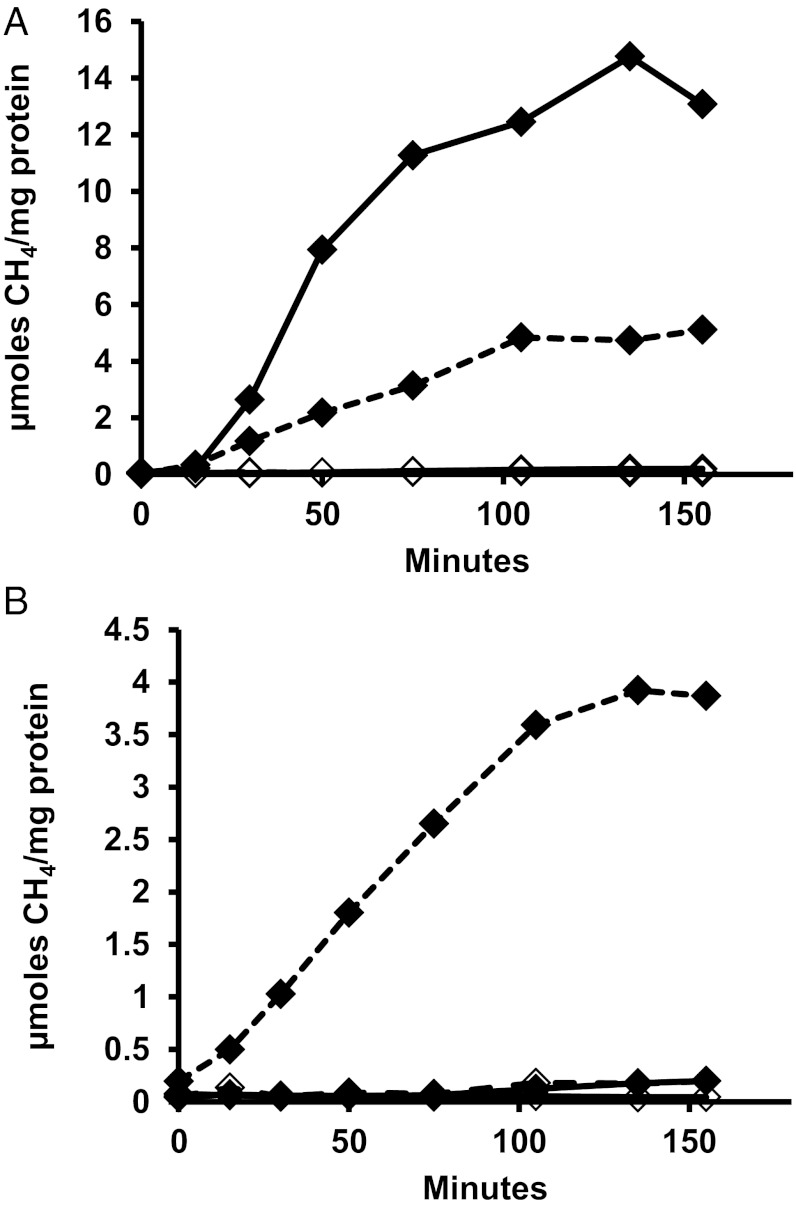
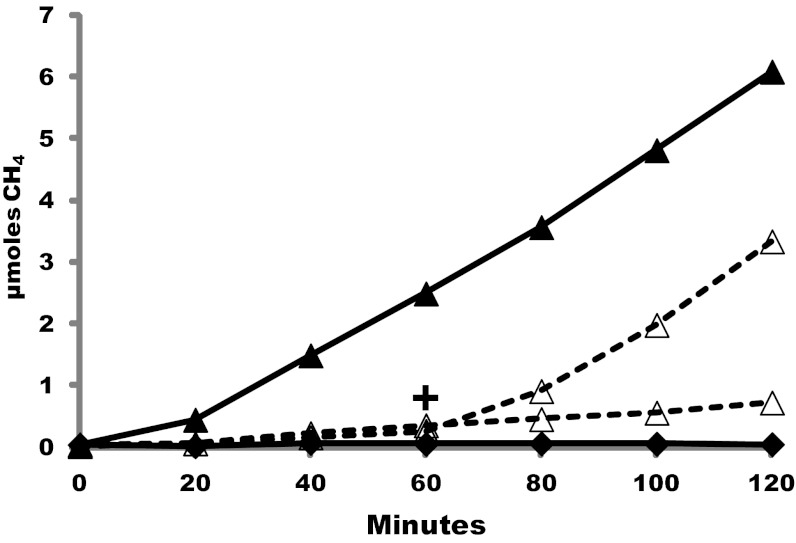
Despite decades of study, electron flow and energy conservation in methanogenic Archaea are still not thoroughly understood. For methanogens without cytochromes, flavin-based electron bifurcation has been proposed as an essential energy-conserving mechanism that couples exergonic and endergonic reactions of methanogenesis. However, an alternative hypothesis posits that the energy-converting hydrogenase Eha provides a chemiosmosis-driven electron input to the endergonic reaction. In vivo evidence for both hypotheses is incomplete. By genetically eliminating all nonessential pathways of H(2) metabolism in the model methanogen Methanococcus maripaludis and using formate as an additional electron donor, we isolate electron flow for methanogenesis from flux through Eha. We find that Eha does not function stoichiometrically for methanogenesis, implying that electron bifurcation must operate in vivo. We show that Eha is nevertheless essential, and a substoichiometric requirement for H(2) suggests that its role is anaplerotic. Indeed, H(2) via Eha stimulates methanogenesis from formate when intermediates are not otherwise replenished. These results fit the model for electron bifurcation, which renders the methanogenic pathway cyclic, and as such requires the replenishment of intermediates. Defining a role for Eha and verifying electron bifurcation provide a complete model of methanogenesis where all necessary electron inputs are accounted for.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
The Wolfe cycle comes full circle.Proc Natl Acad Sci U S A. 2012 Sep 18;109(38):15084-5. doi: 10.1073/pnas.1213193109. Epub 2012 Sep 5. Proc Natl Acad Sci U S A. 2012. PMID: 22955879 Free PMC article. No abstract available.
Similar articles
-
H2-independent growth of the hydrogenotrophic methanogen Methanococcus maripaludis.mBio. 2013 Feb 26;4(2):e00062-13. doi: 10.1128/mBio.00062-13. mBio. 2013. PMID: 23443005 Free PMC article.
-
Protein complexing in a methanogen suggests electron bifurcation and electron delivery from formate to heterodisulfide reductase.Proc Natl Acad Sci U S A. 2010 Jun 15;107(24):11050-5. doi: 10.1073/pnas.1003653107. Epub 2010 Jun 1. Proc Natl Acad Sci U S A. 2010. PMID: 20534465 Free PMC article.
-
Exploring Hydrogenotrophic Methanogenesis: a Genome Scale Metabolic Reconstruction of Methanococcus maripaludis.J Bacteriol. 2016 Nov 18;198(24):3379-3390. doi: 10.1128/JB.00571-16. Print 2016 Dec 15. J Bacteriol. 2016. PMID: 27736793 Free PMC article.
-
Energy Conservation and Hydrogenase Function in Methanogenic Archaea, in Particular the Genus Methanosarcina.Microbiol Mol Biol Rev. 2019 Sep 18;83(4):e00020-19. doi: 10.1128/MMBR.00020-19. Print 2019 Nov 20. Microbiol Mol Biol Rev. 2019. PMID: 31533962 Free PMC article. Review.
-
Electron Bifurcation and Confurcation in Methanogenesis and Reverse Methanogenesis.Front Microbiol. 2018 Jun 20;9:1322. doi: 10.3389/fmicb.2018.01322. eCollection 2018. Front Microbiol. 2018. PMID: 29973922 Free PMC article. Review.
Cited by
-
HydDB: A web tool for hydrogenase classification and analysis.Sci Rep. 2016 Sep 27;6:34212. doi: 10.1038/srep34212. Sci Rep. 2016. PMID: 27670643 Free PMC article.
-
Identification of key components in the energy metabolism of the hyperthermophilic sulfate-reducing archaeon Archaeoglobus fulgidus by transcriptome analyses.Front Microbiol. 2014 Mar 11;5:95. doi: 10.3389/fmicb.2014.00095. eCollection 2014. Front Microbiol. 2014. PMID: 24672515 Free PMC article.
-
Progress of Crude Oil Gasification Technology Assisted by Microorganisms in Reservoirs.Microorganisms. 2024 Mar 29;12(4):702. doi: 10.3390/microorganisms12040702. Microorganisms. 2024. PMID: 38674646 Free PMC article. Review.
-
Hydrogenase-independent uptake and metabolism of electrons by the archaeon Methanococcus maripaludis.ISME J. 2014 Aug;8(8):1673-81. doi: 10.1038/ismej.2014.82. Epub 2014 May 20. ISME J. 2014. PMID: 24844759 Free PMC article.
-
New mode of energy metabolism in the seventh order of methanogens as revealed by comparative genome analysis of “Candidatus methanoplasma termitum”.Appl Environ Microbiol. 2015 Feb;81(4):1338-52. doi: 10.1128/AEM.03389-14. Appl Environ Microbiol. 2015. PMID: 25501486 Free PMC article.
References
-
- Thauer RK, Kaster AK, Seedorf H, Buckel W, Hedderich R. Methanogenic archaea: Ecologically relevant differences in energy conservation. Nat Rev Microbiol. 2008;6:579–591. - PubMed
-
- Hedderich R. Energy-converting [NiFe] hydrogenases from archaea and extremophiles: Ancestors of complex I. J Bioenerg Biomembr. 2004;36:65–75. - PubMed
-
- Gunsalus RP, Wolfe RS. Stimulation of CO2 reduction to methane by methylcoenzyme M in extracts Methanobacterium. Biochem Biophys Res Commun. 1977;76:790–795. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
