

Cytoskeleton assembly in Toxoplasma gondii cell division
- PMID: 22878103
- PMCID: PMC4066374
- DOI: 10.1016/B978-0-12-394309-5.00001-8
Cytoskeleton assembly in Toxoplasma gondii cell division
Abstract
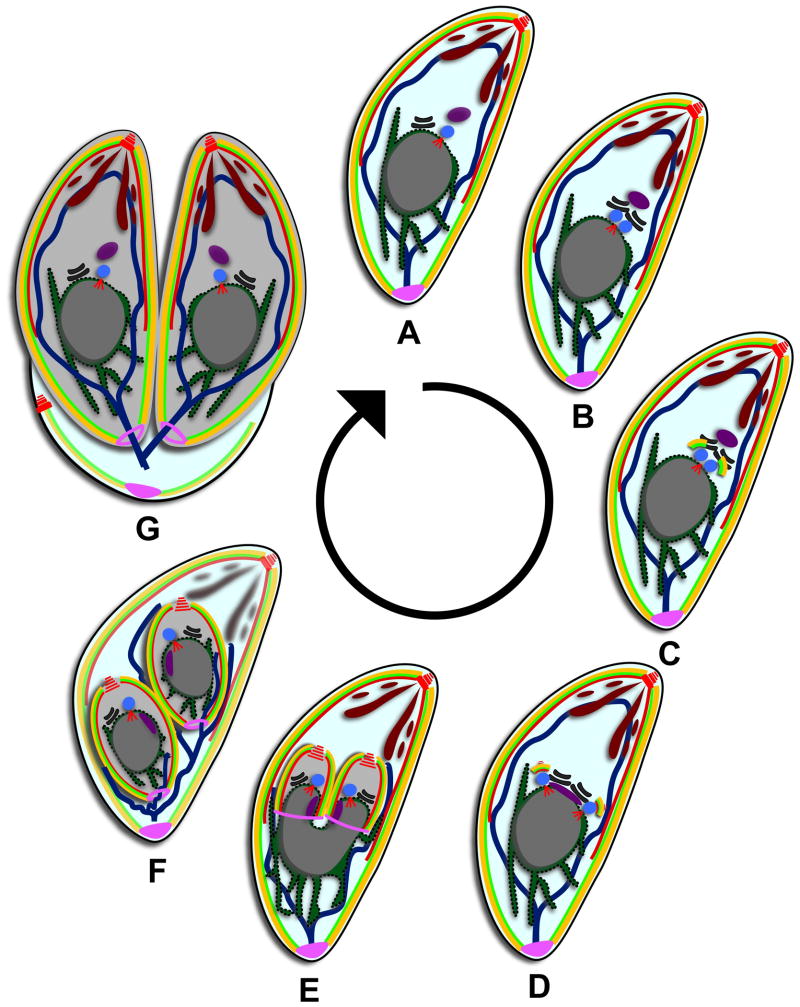
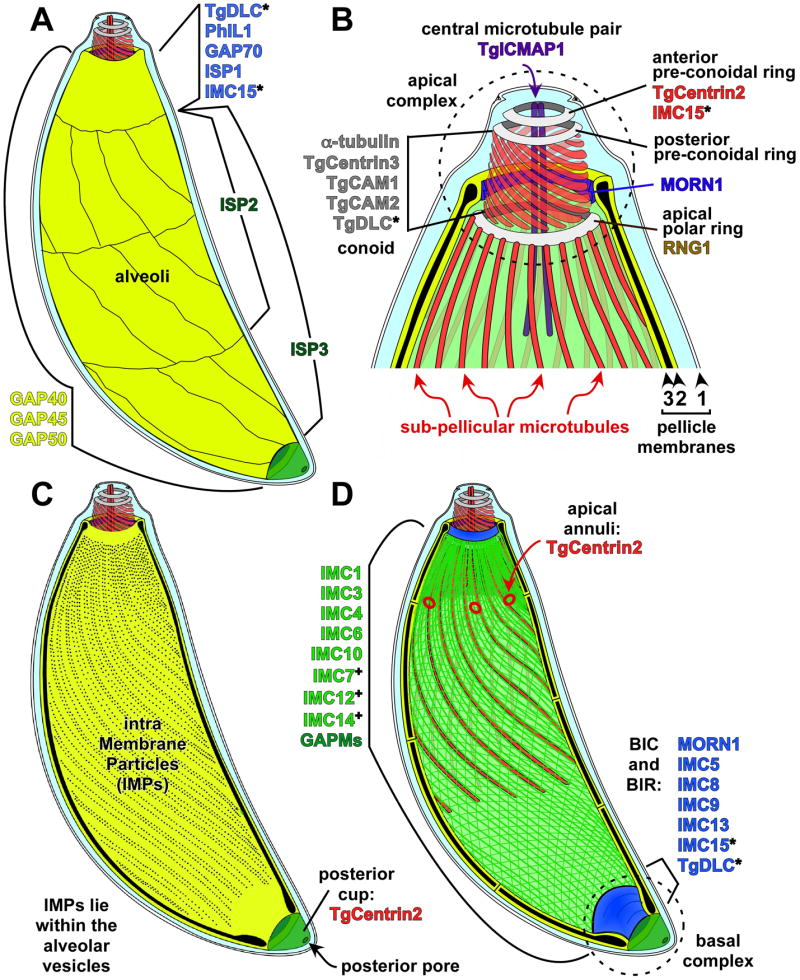
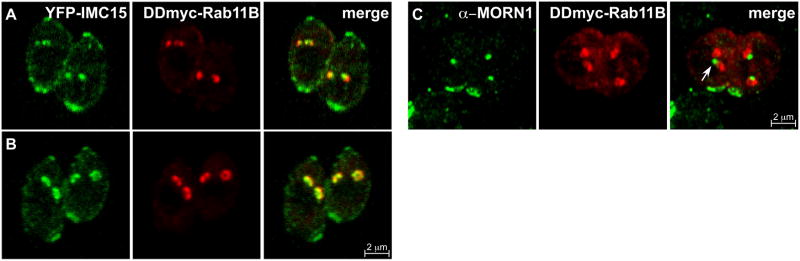
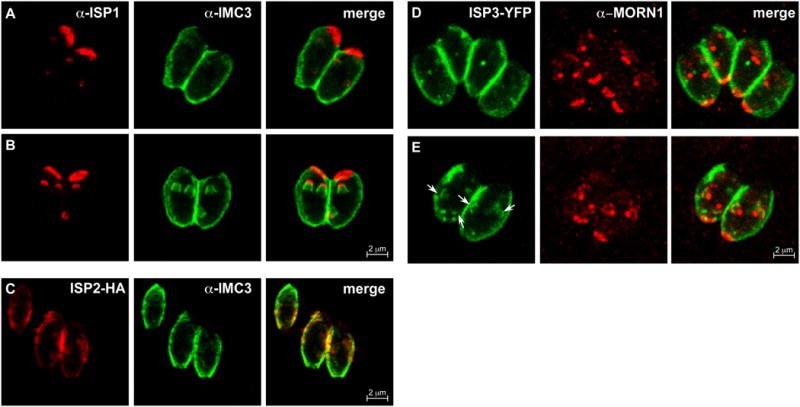
Cell division across members of the protozoan parasite phylum Apicomplexa displays a surprising diversity between different species as well as between different life stages of the same parasite. In most cases, infection of a host cell by a single parasite results in the formation of a polyploid cell from which individual daughters bud in a process dependent on a final round of mitosis. Unlike other apicomplexans, Toxoplasma gondii divides by a binary process consisting of internal budding that results in only two daughter cells per round of division. Since T. gondii is experimentally accessible and displays the simplest division mode, it has manifested itself as a model for apicomplexan daughter formation. Here, we review newly emerging insights in the prominent role that assembly of the cortical cytoskeletal scaffold plays in the process of daughter parasite formation.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
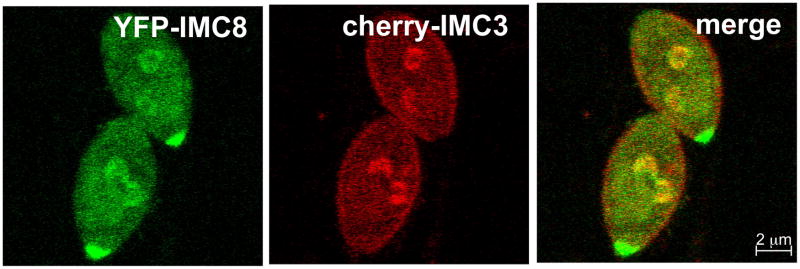
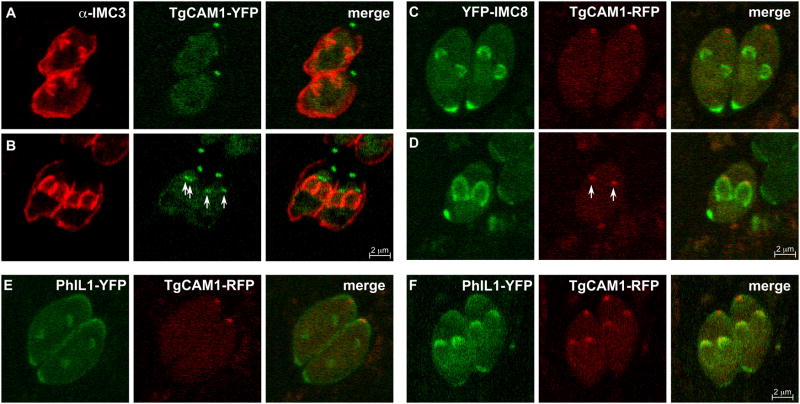
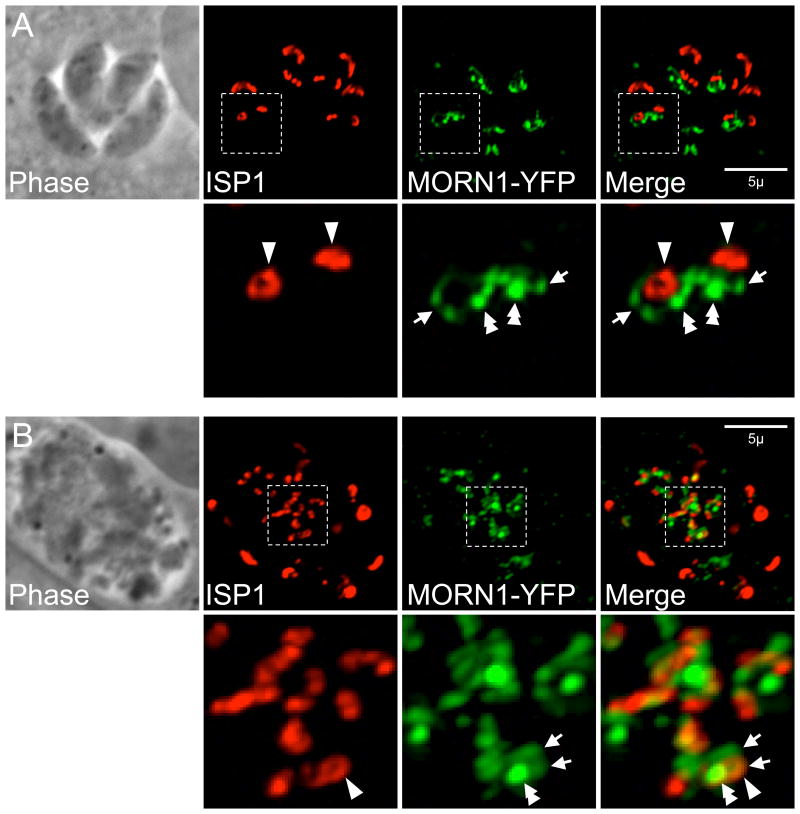
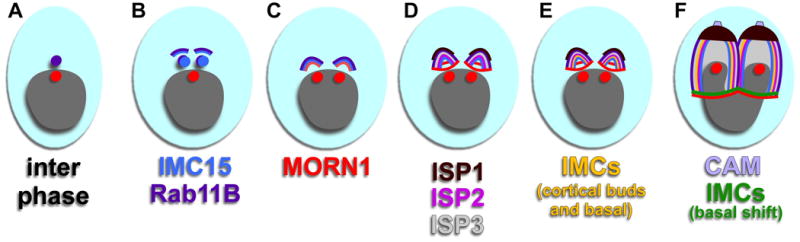
Similar articles
-
Daughter cell assembly in the protozoan parasite Toxoplasma gondii.Mol Biol Cell. 2002 Feb;13(2):593-606. doi: 10.1091/mbc.01-06-0309. Mol Biol Cell. 2002. PMID: 11854415 Free PMC article.
-
Organizational changes of the daughter basal complex during the parasite replication of Toxoplasma gondii.PLoS Pathog. 2008 Jan;4(1):e10. doi: 10.1371/journal.ppat.0040010. PLoS Pathog. 2008. PMID: 18208326 Free PMC article.
-
Fussing About Fission: Defining Variety Among Mainstream and Exotic Apicomplexan Cell Division Modes.Front Cell Infect Microbiol. 2020 Jun 5;10:269. doi: 10.3389/fcimb.2020.00269. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32582569 Free PMC article.
-
The Riveting Cellular Structures of Apicomplexan Parasites.Trends Parasitol. 2020 Dec;36(12):979-991. doi: 10.1016/j.pt.2020.09.001. Epub 2020 Sep 30. Trends Parasitol. 2020. PMID: 33011071 Review.
-
The cell cycle and Toxoplasma gondii cell division: tightly knit or loosely stitched?Int J Parasitol. 2008 Oct;38(12):1343-58. doi: 10.1016/j.ijpara.2008.06.004. Epub 2008 Jul 24. Int J Parasitol. 2008. PMID: 18703066 Review.
Cited by
-
Apical annuli are specialised sites of post-invasion secretion of dense granules in Toxoplasma.Elife. 2024 Jan 25;13:e94201. doi: 10.7554/eLife.94201. Elife. 2024. PMID: 38270431 Free PMC article.
-
The Toxoplasma protein phosphatase 6 catalytic subunit (TgPP6C) is essential for cell cycle progression and virulence.PLoS Pathog. 2023 Dec 13;19(12):e1011831. doi: 10.1371/journal.ppat.1011831. eCollection 2023 Dec. PLoS Pathog. 2023. PMID: 38091362 Free PMC article.
-
TgTKL4 Is a Novel Kinase That Plays an Important Role in Toxoplasma Morphology and Fitness.mSphere. 2023 Apr 20;8(2):e0064922. doi: 10.1128/msphere.00649-22. Epub 2023 Feb 14. mSphere. 2023. PMID: 36786615 Free PMC article.
-
Requirement of Toxoplasma gondii metacaspases for IMC1 maturation, endodyogeny and virulence in mice.Parasit Vectors. 2021 Aug 12;14(1):400. doi: 10.1186/s13071-021-04878-0. Parasit Vectors. 2021. PMID: 34384491 Free PMC article.
-
Co-dependent formation of the Toxoplasma gondii sub-pellicular microtubules and inner membrane skeleton.bioRxiv [Preprint]. 2024 May 25:2024.05.25.595886. doi: 10.1101/2024.05.25.595886. bioRxiv. 2024. Update in: mBio. 2025 Aug 13:e0138925. doi: 10.1128/mbio.01389-25. PMID: 38826480 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
