

Aging of the microenvironment influences clonality in hematopoiesis
- PMID: 22879906
- PMCID: PMC3412859
- DOI: 10.1371/journal.pone.0042080
Aging of the microenvironment influences clonality in hematopoiesis
Abstract
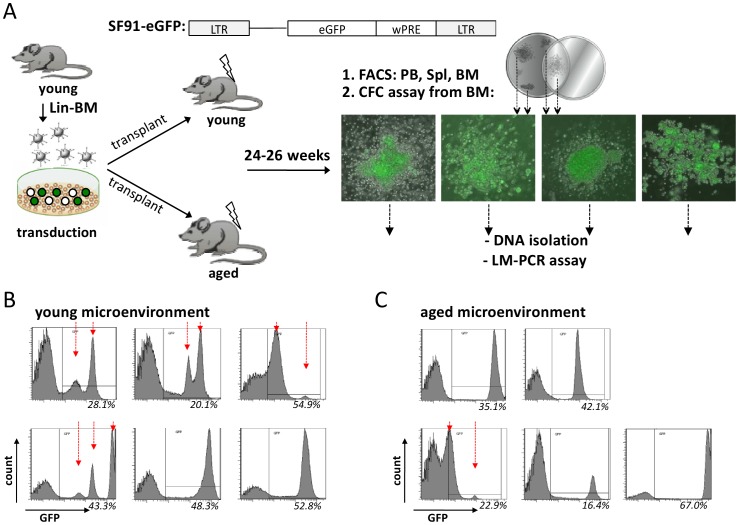
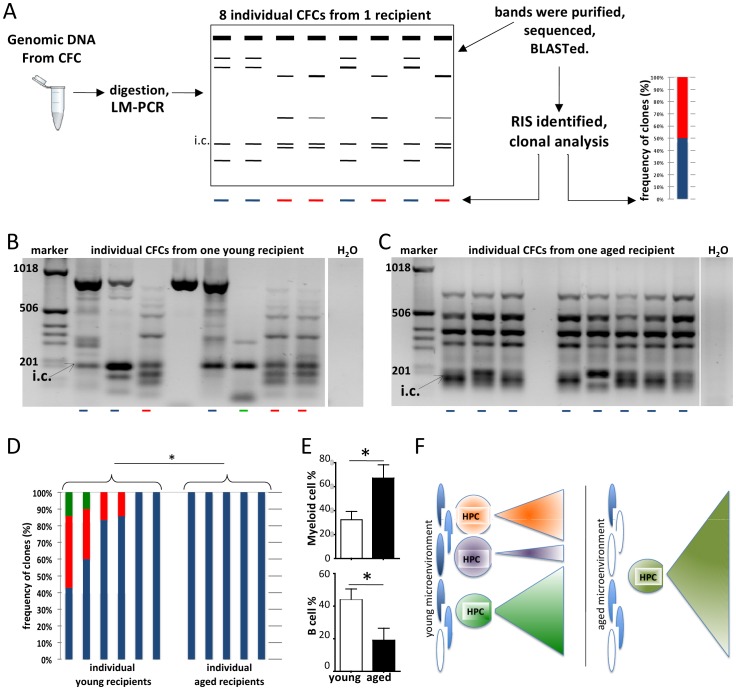
The mechanisms of the age-associated exponential increase in the incidence of leukemia are not known in detail. Leukemia as well as aging are initiated and regulated in multi-factorial fashion by cell-intrinsic and extrinsic factors. The role of aging of the microenvironment for leukemia initiation/progression has not been investigated in great detail so far. Clonality in hematopoiesis is tightly linked to the initiation of leukemia. Based on a retroviral-insertion mutagenesis approach to generate primitive hematopoietic cells with an intrinsic potential for clonal expansion, we determined clonality of transduced hematopoietic progenitor cells (HPCs) exposed to a young or aged microenvironment in vivo. While HPCs displayed primarily oligo-clonality within a young microenvironment, aged animals transplanted with identical pool of cells displayed reduced clonality within transduced HPCs. Our data show that an aged niche exerts a distinct selection pressure on dominant HPC-clones thus facilitating the transition to mono-clonality, which might be one underlying cause for the increased age-associated incidence of leukemia.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
