

fussel (fuss)--A negative regulator of BMP signaling in Drosophila melanogaster
- PMID: 22879948
- PMCID: PMC3413677
- DOI: 10.1371/journal.pone.0042349
fussel (fuss)--A negative regulator of BMP signaling in Drosophila melanogaster
Abstract
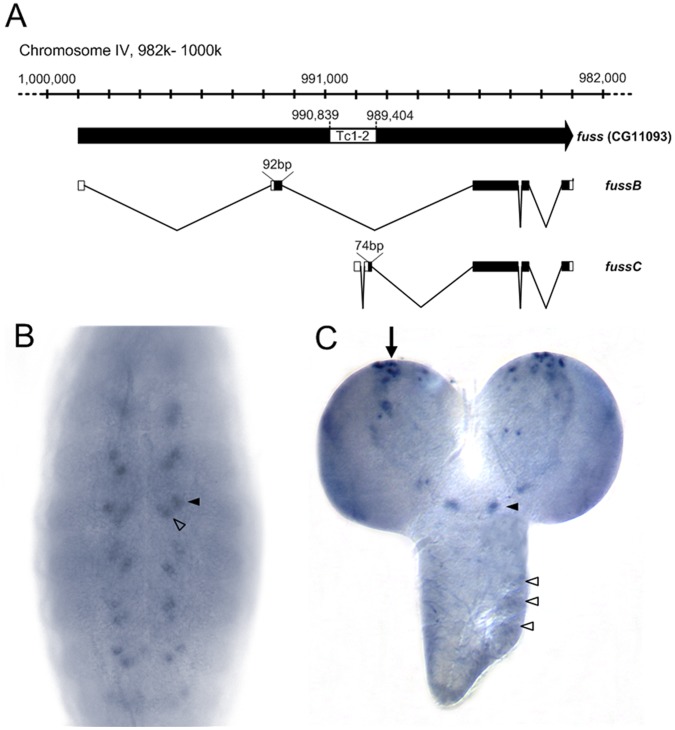
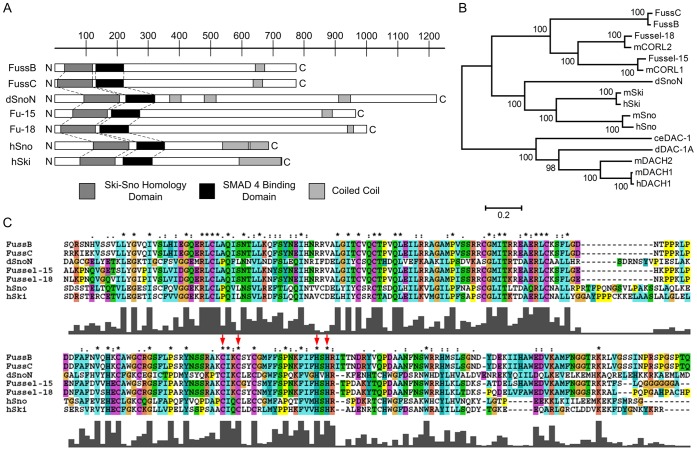
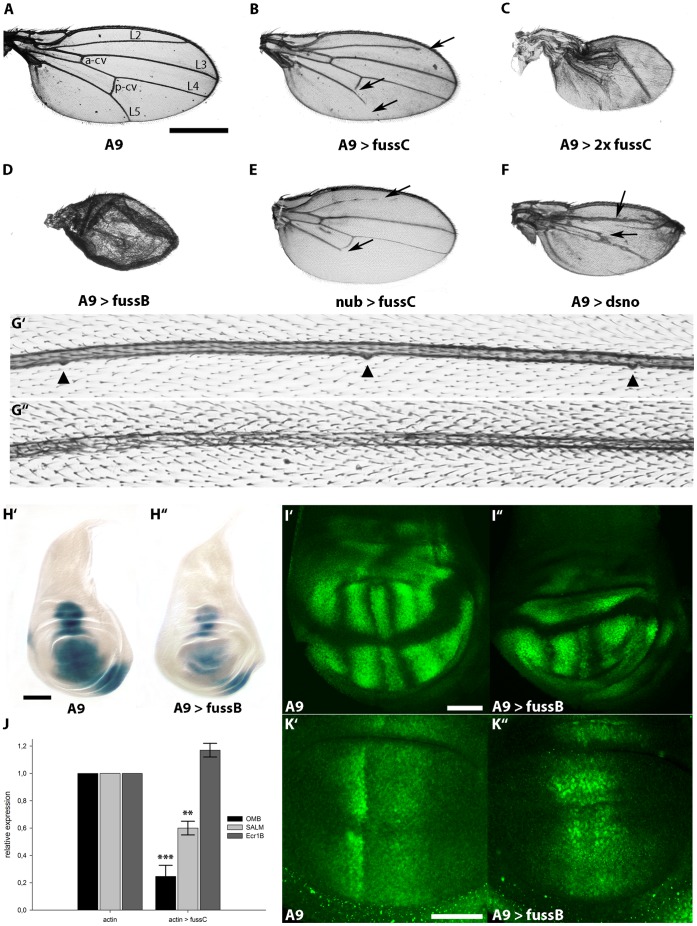
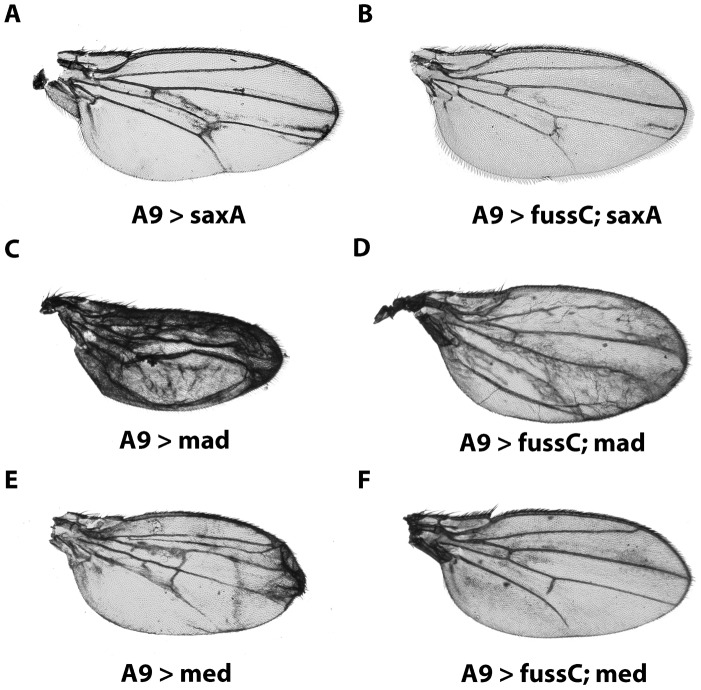
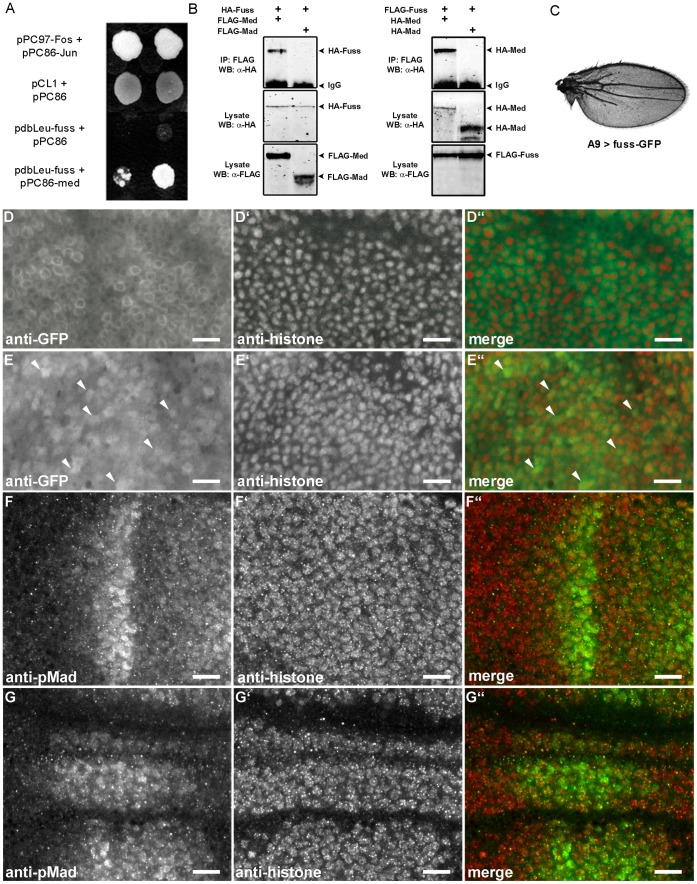
The TGF-β/BMP signaling cascades control a wide range of developmental and physiological functions in vertebrates and invertebrates. In Drosophila melanogaster, members of this pathway can be divided into a Bone Morphogenic Protein (BMP) and an Activin-ß (Act-ß) branch, where Decapentaplegic (Dpp), a member of the BMP family has been most intensively studied. They differ in ligands, receptors and transmitting proteins, but also share some components, such as the Co-Smad Medea (Med). The essential role of Med is to form a complex with one of the two activating Smads, mothers against decapentaplegic (Mad) or dSmad, and to translocate together to the nucleus where they can function as transcriptional regulators of downstream target genes. This signaling cascade underlies different mechanisms of negative regulation, which can be exerted by inhibitory Smads, such as daughters against decapentaplegic (dad), but also by the Ski-Sno family. In this work we identified and functionally analyzed a new member of the Ski/Sno-family, fussel (fuss), the Drosophila homolog of the human functional suppressing element 15 (fussel-15). fuss codes for two differentially spliced transcripts with a neuronal expression pattern. The proteins are characterized by a Ski-Sno and a SAND homology domain. Overexpression studies and genetic interaction experiments clearly reveal an interaction of fuss with members of the BMP pathway, leading to a strong repression of BMP-signaling. The protein interacts directly with Medea and seems to reprogram the Smad pathway through its influence upon the formation of functional Mad/Medea complexes. This leads amongst others to a repression of downstream target genes of the Dpp pathway, such as optomotor blind (omb). Taken together we could show that fuss exerts a pivotal role as an antagonist of BMP signaling in Drosophila melanogaster.
Conflict of interest statement
Figures
References
-
- Padgett RW, St Johnston RD, Gelbart WM (1987) A transcript from a Drosophila pattern gene predicts a protein homologous to the transforming growth factor-beta family. Nature 325: 81–84. - PubMed
-
- Wozney JM, Rosen V, Celeste AJ, Mitsock LM, Whitters MJ, et al. (1988) Novel regulators of bone formation: molecular clones and activities. Science 242: 1528–1534. - PubMed
-
- Twombly V, Blackman RK, Jin H, Graff JM, Padgett RW, et al. (1996) The TGF-beta signaling pathway is essential for Drosophila oogenesis. Development 122: 1555–1565. - PubMed
-
- Shravage BV, Altmann G, Technau M, Roth S (2007) The role of Dpp and its inhibitors during eggshell patterning in Drosophila . Development 134: 2261–2271. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
