

Widespread state-dependent shifts in cerebellar activity in locomoting mice
- PMID: 22880068
- PMCID: PMC3411825
- DOI: 10.1371/journal.pone.0042650
Widespread state-dependent shifts in cerebellar activity in locomoting mice
Abstract
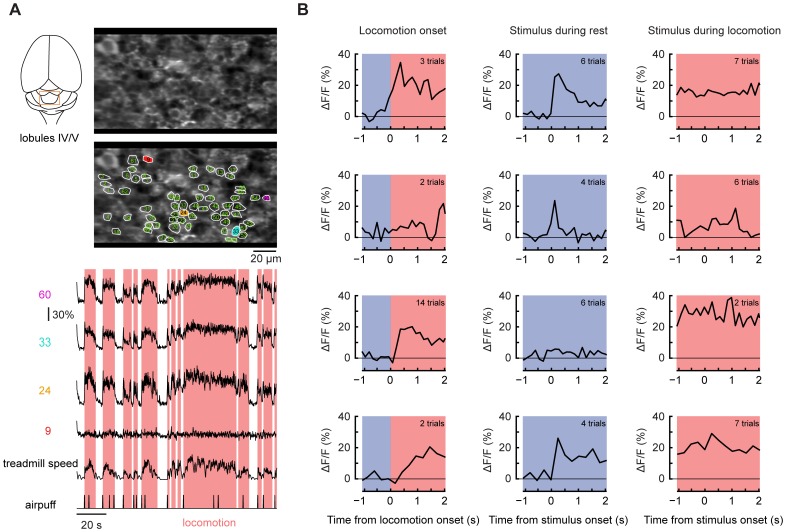
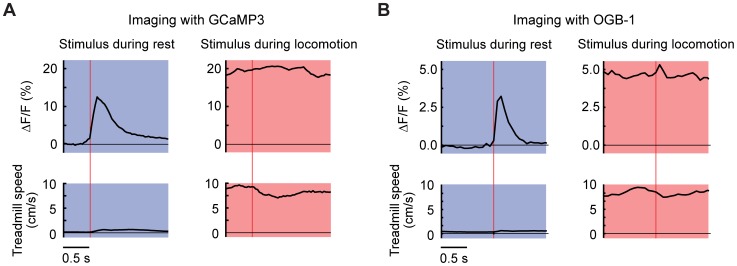
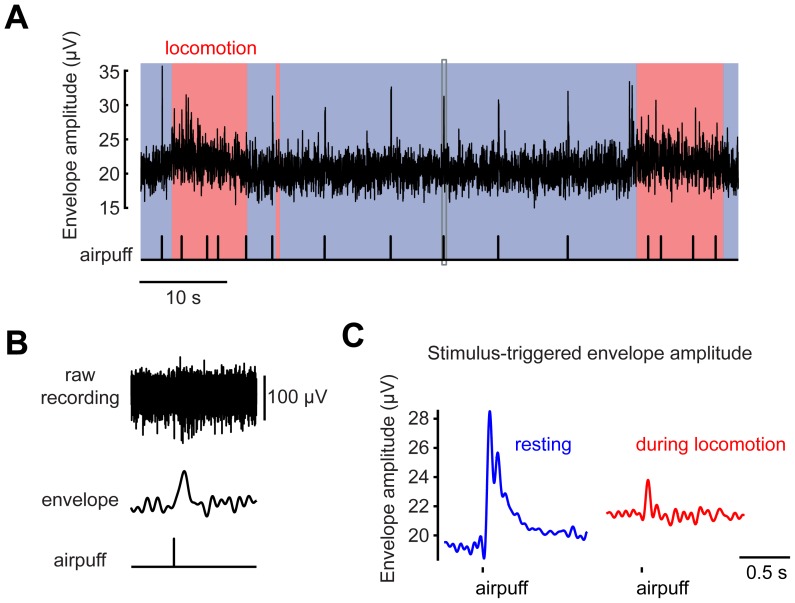
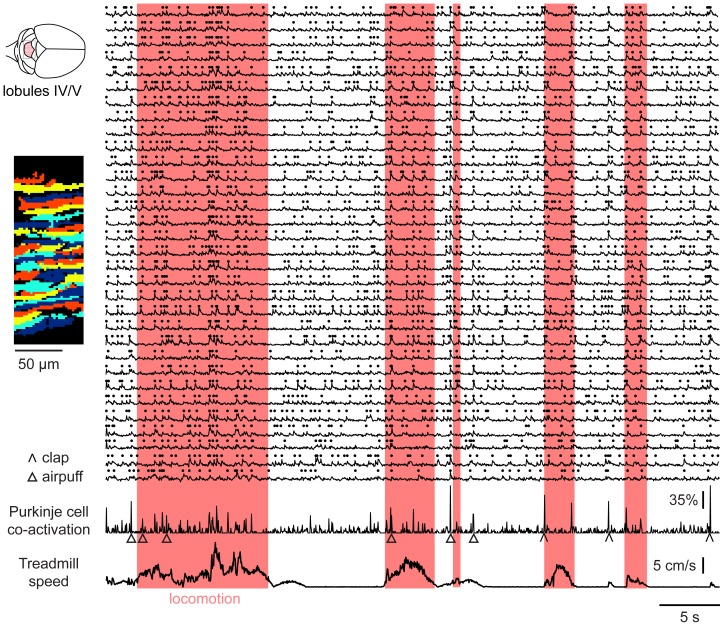
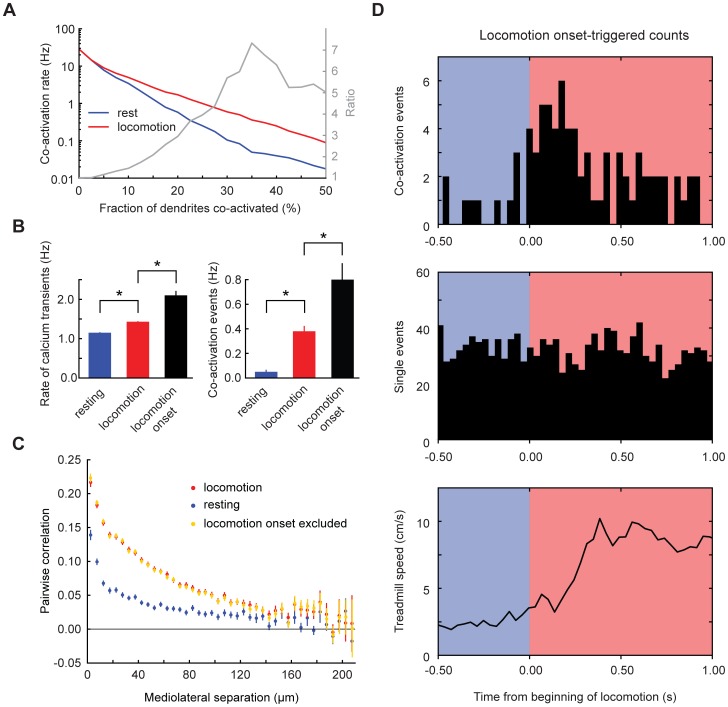
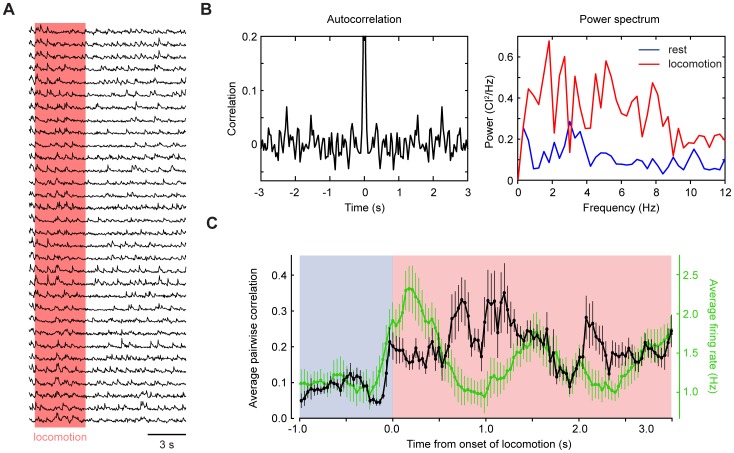
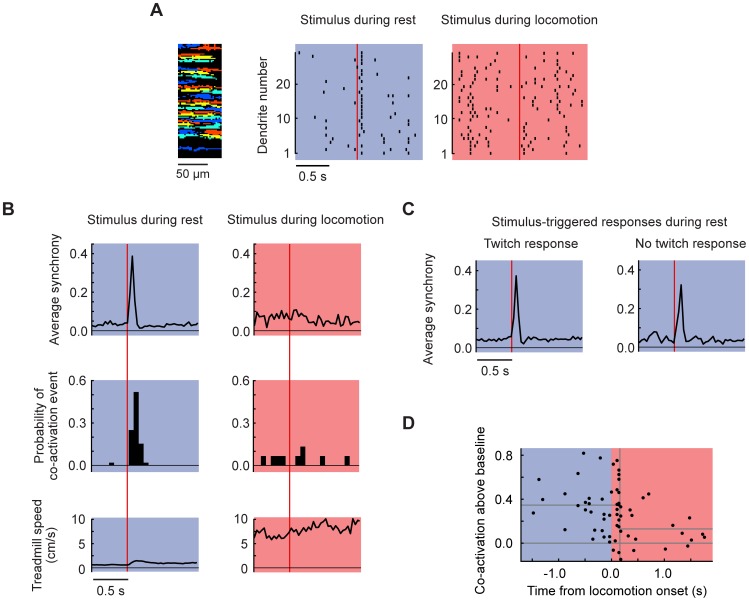
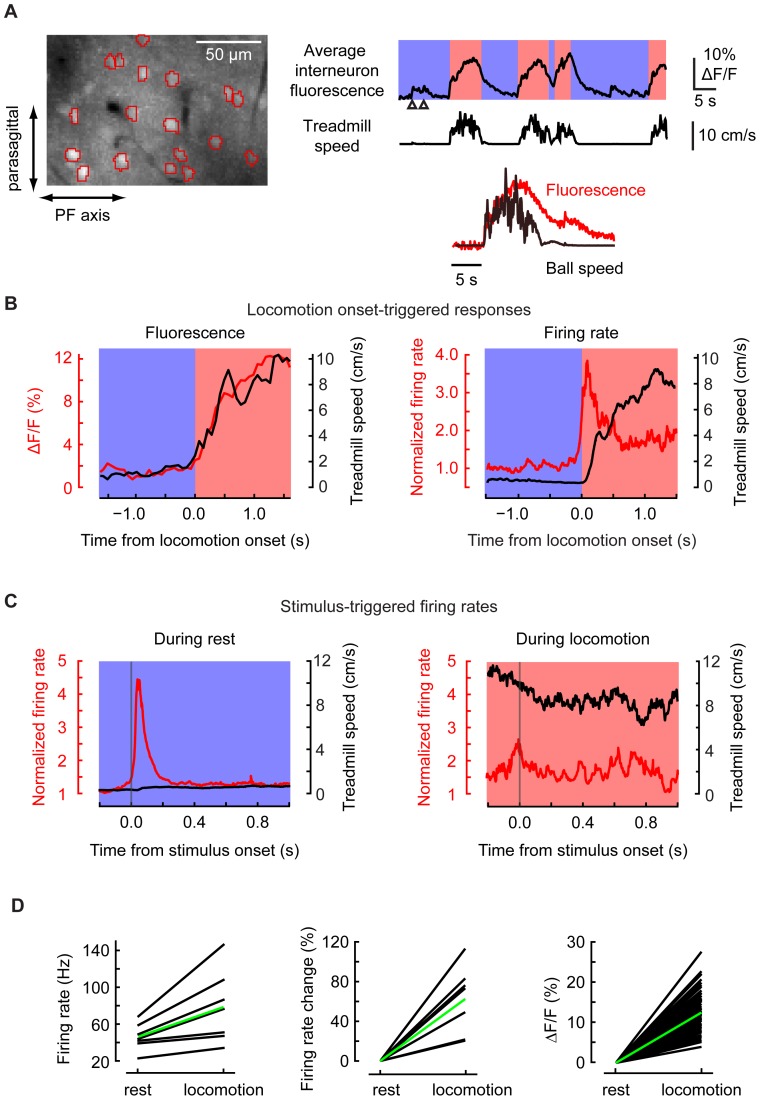
Excitatory drive enters the cerebellum via mossy fibers, which activate granule cells, and climbing fibers, which activate Purkinje cell dendrites. Until now, the coordinated regulation of these pathways has gone unmonitored in spatially resolved neuronal ensembles, especially in awake animals. We imaged cerebellar activity using functional two-photon microscopy and extracellular recording in awake mice locomoting on an air-cushioned spherical treadmill. We recorded from putative granule cells, molecular layer interneurons, and Purkinje cell dendrites in zone A of lobule IV/V, representing sensation and movement from trunk and limbs. Locomotion was associated with widespread increased activity in granule cells and interneurons, consistent with an increase in mossy fiber drive. At the same time, dendrites of different Purkinje cells showed increased co-activation, reflecting increased synchrony of climbing fiber activity. In resting animals, aversive stimuli triggered increased activity in granule cells and interneurons, as well as increased Purkinje cell co-activation that was strongest for neighboring dendrites and decreased smoothly as a function of mediolateral distance. In contrast with anesthetized recordings, no 1-10 Hz oscillations in climbing fiber activity were evident. Once locomotion began, responses to external stimuli in all three cell types were strongly suppressed. Thus climbing and mossy fiber representations can shift together within a fraction of a second, reflecting in turn either movement-associated activity or external stimuli.
Conflict of interest statement
Figures
References
-
- Bell CC, Han V, Sawtell NB (2008) Cerebellum-like structures and their implications for cerebellar function. Annu Rev Neurosci 31: 1–24. - PubMed
-
- Bloedel JR, Bracha V (1998) Current concepts of climbing fiber function. Anat Rec 253: 118–126. - PubMed
-
- Eccles JC, Provini L, Strata P, Táboříková H (1968) Analysis of electrical potentials evoked in the cerebellar anterior lobe by stimulation of hindlimb and forelimb nerves. Exp Brain Res 6: 171–194. - PubMed
-
- Eccles JC, Faber DS, Murphy JT, Sabah NH, Táboříková H (1971) Afferent volleys in limb nerves influencing impulse discharges in cerebellar cortex. I. In mossy fibers and granule cells. Exp Brain Res 13: 15–35. - PubMed
-
- Chadderton P, Margrie TW, Häusser M (2004) Integration of quanta in cerebellar granule cells during sensory processing. Nature 428: 856–860. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
