

Burkitt lymphoma pathogenesis and therapeutic targets from structural and functional genomics
- PMID: 22885699
- PMCID: PMC3609867
- DOI: 10.1038/nature11378
Burkitt lymphoma pathogenesis and therapeutic targets from structural and functional genomics
Abstract
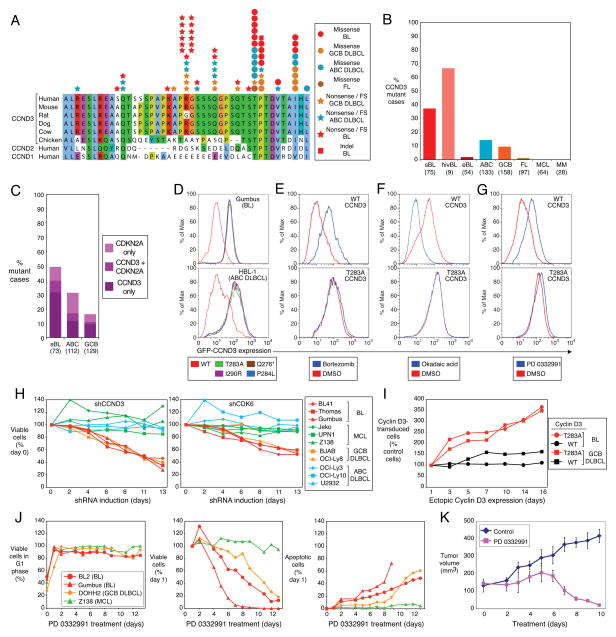
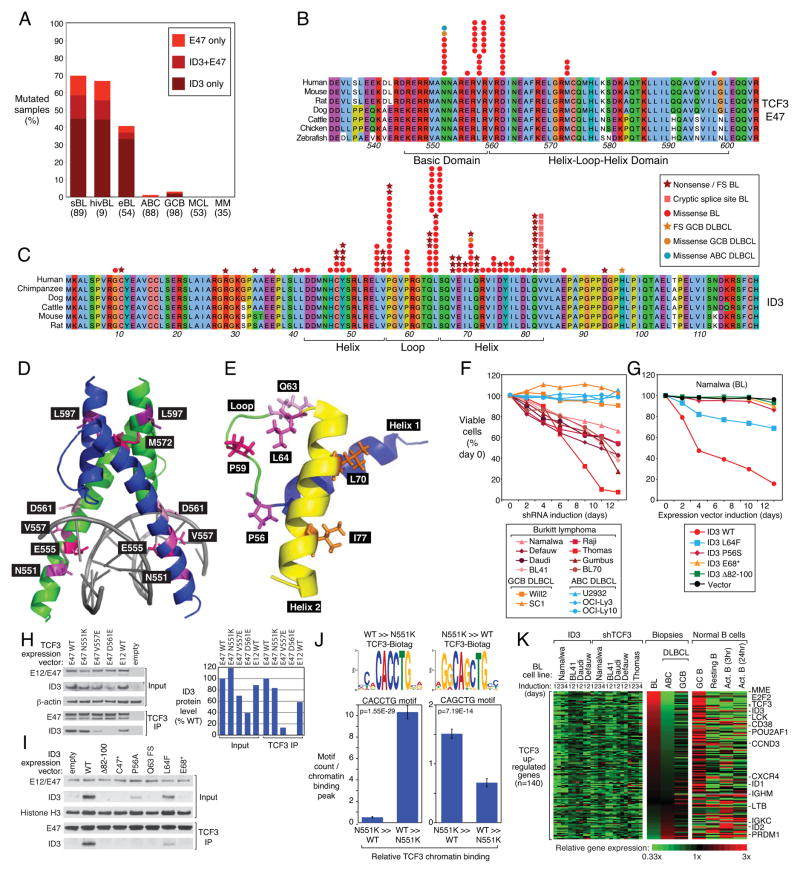
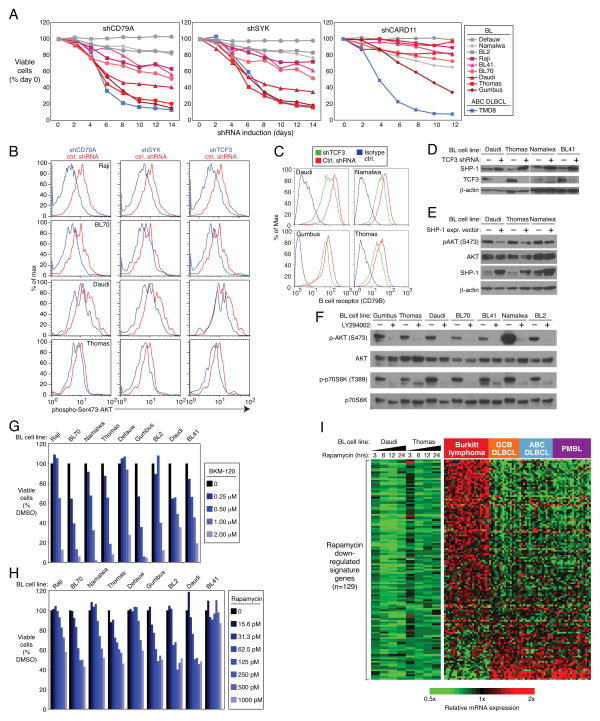
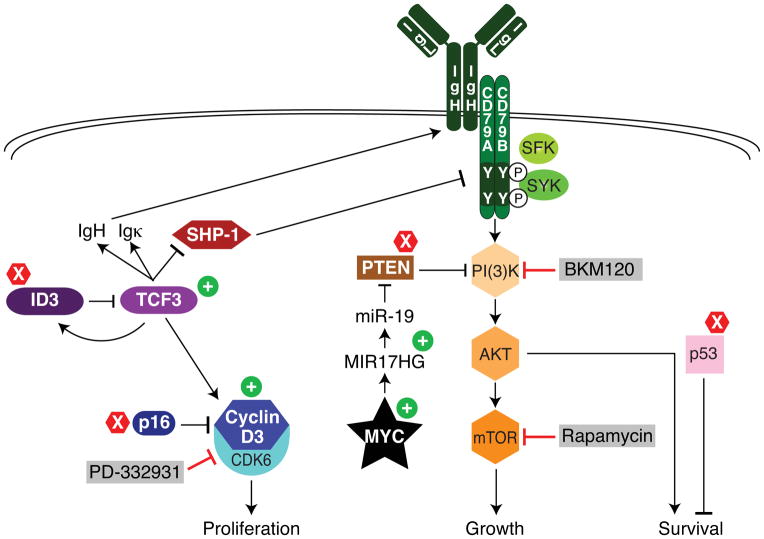
Burkitt's lymphoma (BL) can often be cured by intensive chemotherapy, but the toxicity of such therapy precludes its use in the elderly and in patients with endemic BL in developing countries, necessitating new strategies. The normal germinal centre B cell is the presumed cell of origin for both BL and diffuse large B-cell lymphoma (DLBCL), yet gene expression analysis suggests that these malignancies may use different oncogenic pathways. BL is subdivided into a sporadic subtype that is diagnosed in developed countries, the Epstein-Barr-virus-associated endemic subtype, and an HIV-associated subtype, but it is unclear whether these subtypes use similar or divergent oncogenic mechanisms. Here we used high-throughput RNA sequencing and RNA interference screening to discover essential regulatory pathways in BL that cooperate with MYC, the defining oncogene of this cancer. In 70% of sporadic BL cases, mutations affecting the transcription factor TCF3 (E2A) or its negative regulator ID3 fostered TCF3 dependency. TCF3 activated the pro-survival phosphatidylinositol-3-OH kinase pathway in BL, in part by augmenting tonic B-cell receptor signalling. In 38% of sporadic BL cases, oncogenic CCND3 mutations produced highly stable cyclin D3 isoforms that drive cell cycle progression. These findings suggest opportunities to improve therapy for patients with BL.
Figures
Comment in
-
Targeted therapies: New targets in Burkitt lymphoma?Nat Rev Clin Oncol. 2012 Oct;9(10):551. doi: 10.1038/nrclinonc.2012.153. Epub 2012 Aug 28. Nat Rev Clin Oncol. 2012. PMID: 22926023 No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
