

Mesenchymal stem cells transmigrate between and directly through tumor necrosis factor-α-activated endothelial cells via both leukocyte-like and novel mechanisms
- PMID: 22887987
- PMCID: PMC3479371
- DOI: 10.1002/stem.1198
Mesenchymal stem cells transmigrate between and directly through tumor necrosis factor-α-activated endothelial cells via both leukocyte-like and novel mechanisms
Abstract
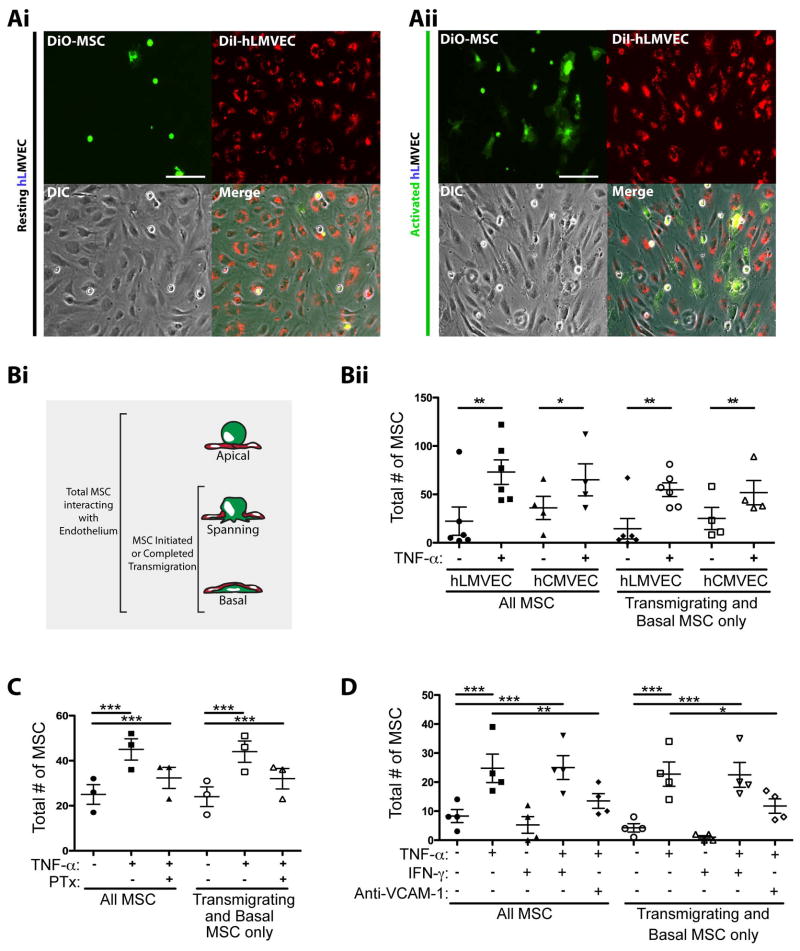
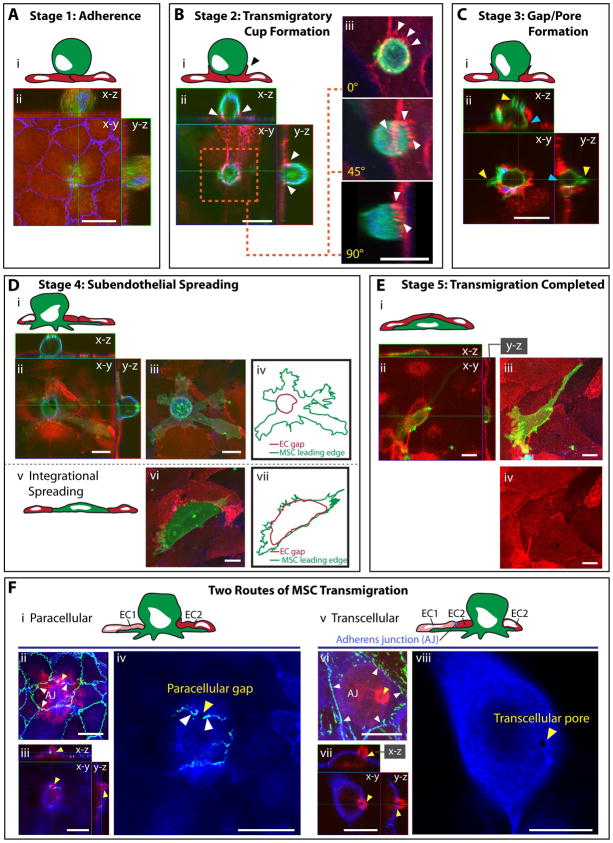
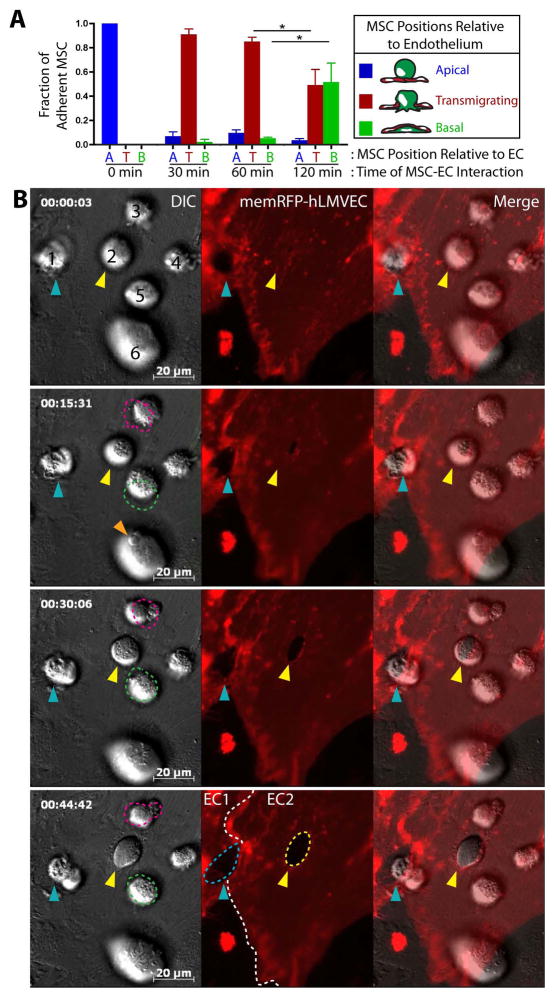
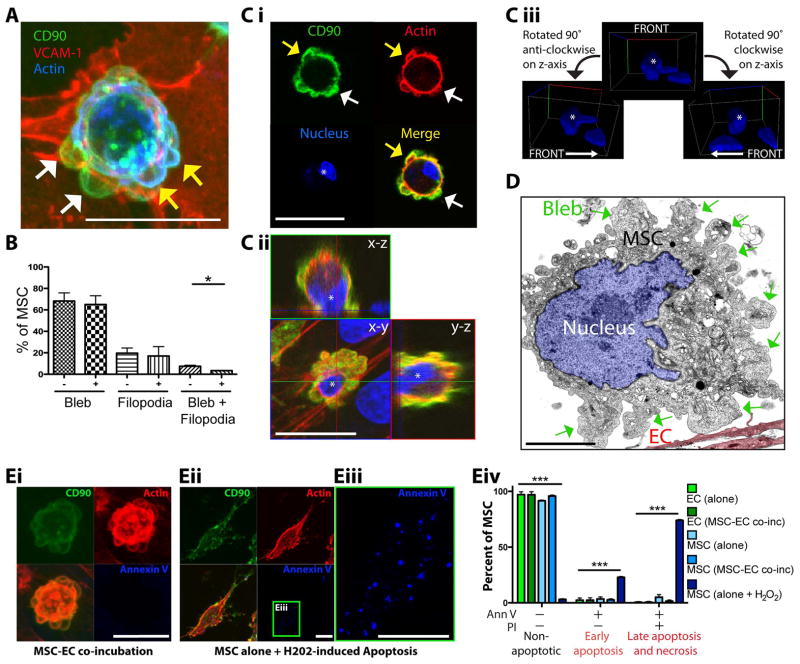
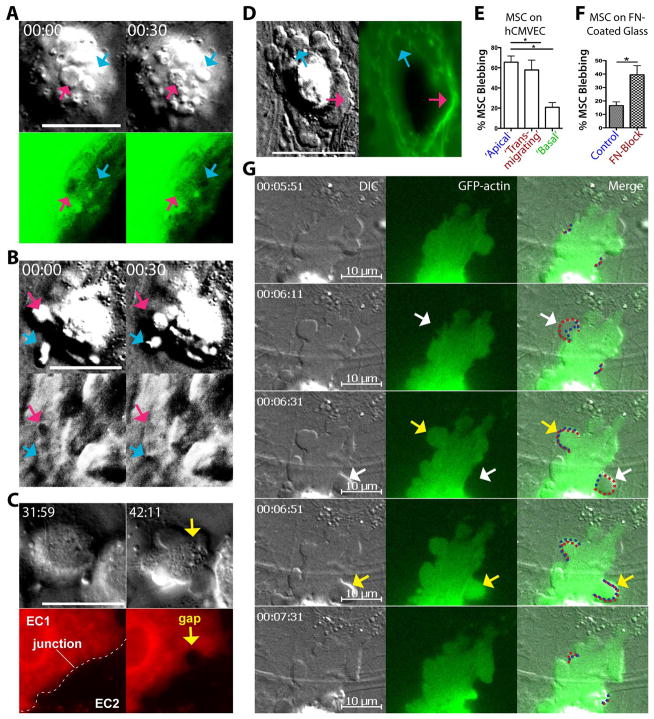
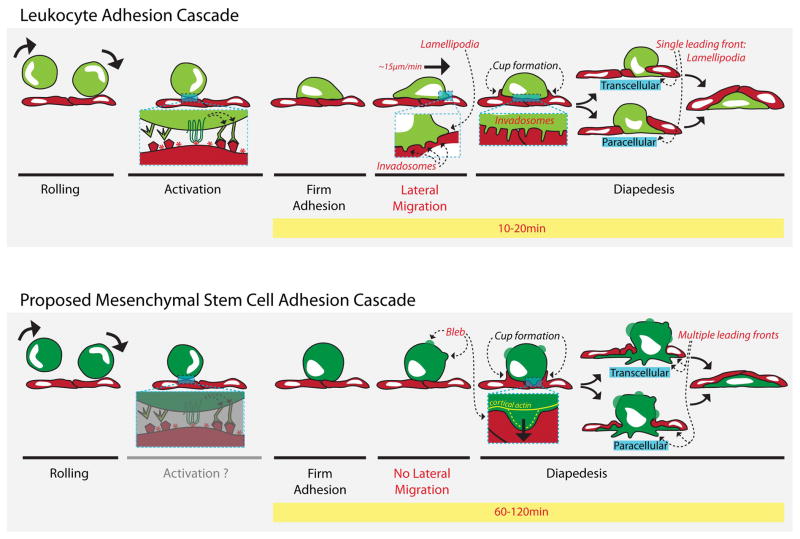
Systemically administered adult mesenchymal stem cells (MSCs), which are being explored in clinical trials to treat inflammatory disease, exhibit the critical ability to extravasate at sites of inflammation. We aimed to characterize the basic cellular processes mediating this extravasation and compare them to those involved in leukocyte transmigration. Using high-resolution confocal and dynamic microscopy, we show that, like leukocytes, human bone marrow-derived MSC preferentially adhere to and migrate across tumor necrosis factor-α-activated endothelium in a vascular cell adhesion molecule-1 (VCAM-1) and G-protein-coupled receptor signaling-dependent manner. As several studies have suggested, we observed that a fraction of MSC was integrated into endothelium. In addition, we observed two modes of transmigration not previously observed for MSC: Paracellular (between endothelial cells) and transcellular (directly through individual endothelial cells) diapedesis through discrete gaps and pores in the endothelial monolayer, in association with VCAM-1-enriched "transmigratory cups". Contrasting leukocytes, MSC transmigration was not preceded by significant lateral migration and occurred on the time scale of hours rather than minutes. Interestingly, rather than lamellipodia and invadosomes, MSC exhibited nonapoptotic membrane blebbing activity that was similar to activities previously described for metastatic tumor and embryonic germ cells. Our studies suggest that low avidity binding between endothelium and MSC may grant a permissive environment for MSC blebbing. MSC blebbing was associated with early stages of transmigration, in which blebs could exert forces on underlying endothelial cells indicating potential functioning in breaching the endothelium. Collectively, our data suggest that MSC transmigrate actively into inflamed tissues via both leukocyte-like and novel mechanisms.
Copyright © 2012 AlphaMed Press.
Figures
References
-
- Karp JM, Teo GSL. Mesenchymal Stem Cell Homing: The Devil is in the Details. Cell Stem Cell. 2009;4:206–216. - PubMed
-
- Kawada H, Fujita J, Kinjo K, et al. Nonhematopoietic mesenchymal stem cells can be mobilized and differentiate into cardiomyocytes after myocardial infarction. Blood. 2004;104:3581–3587. - PubMed
-
- Taylor HS. Endometrial Cells Derived From Donor Stem Cells in Bone Marrow Transplant Recipients. JAMA: The Journal of the American Medical Association. 2004;292:81–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
