

Molecular characterization of banana NAC transcription factors and their interactions with ethylene signalling component EIL during fruit ripening
- PMID: 22888129
- PMCID: PMC3430993
- DOI: 10.1093/jxb/ers178
Molecular characterization of banana NAC transcription factors and their interactions with ethylene signalling component EIL during fruit ripening
Abstract
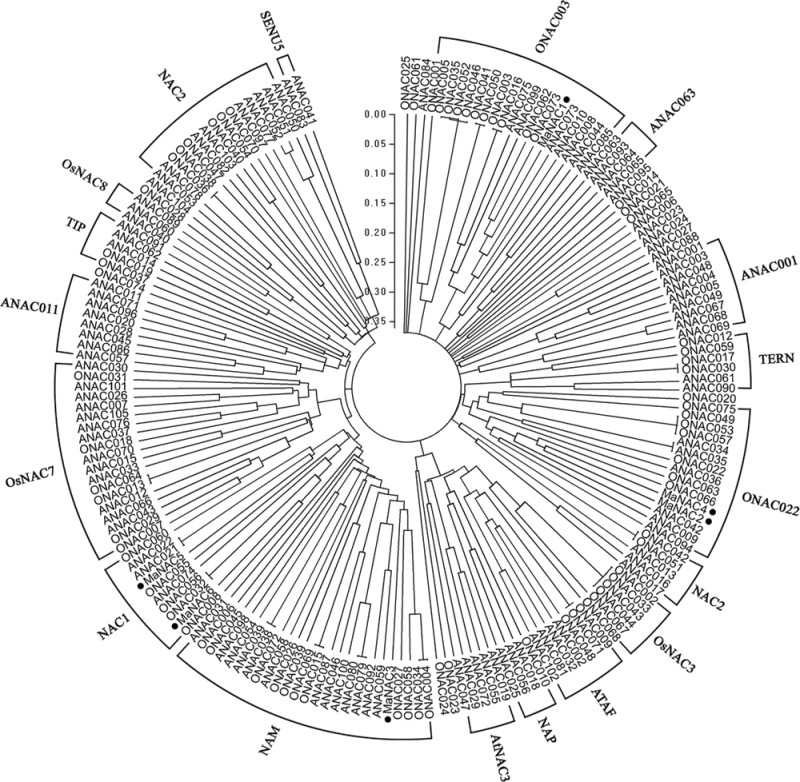
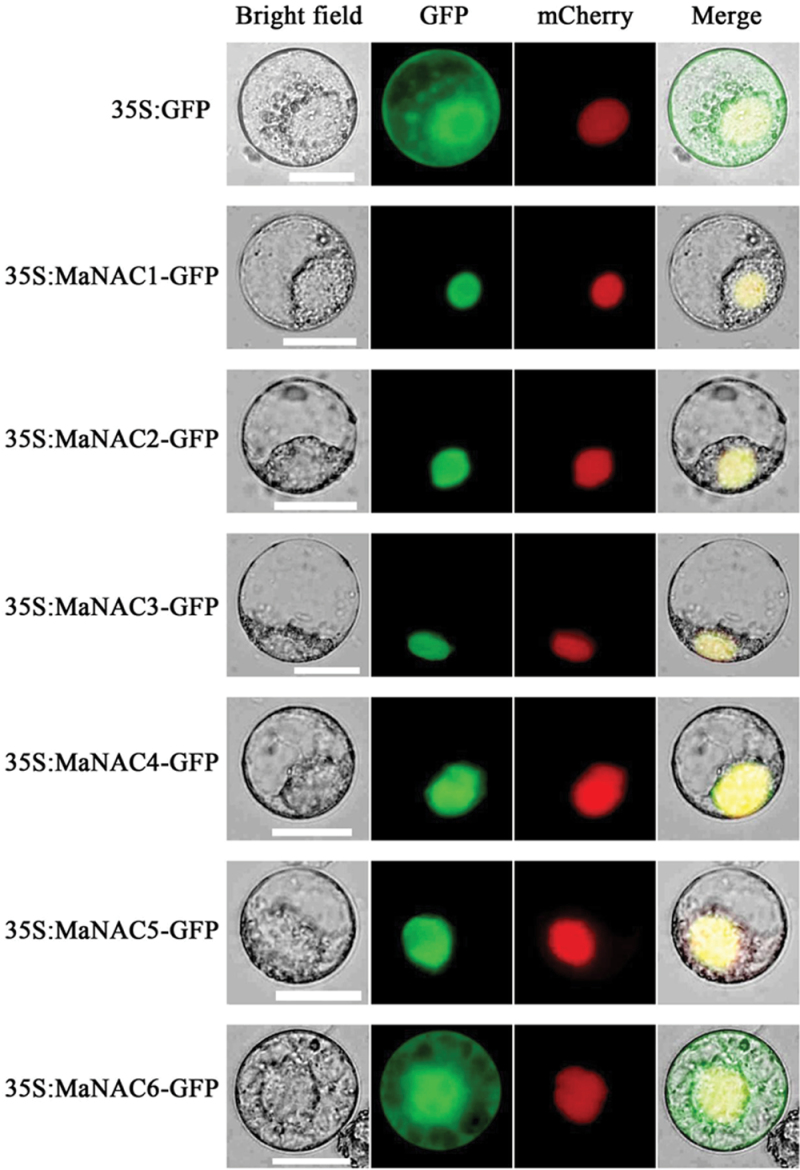
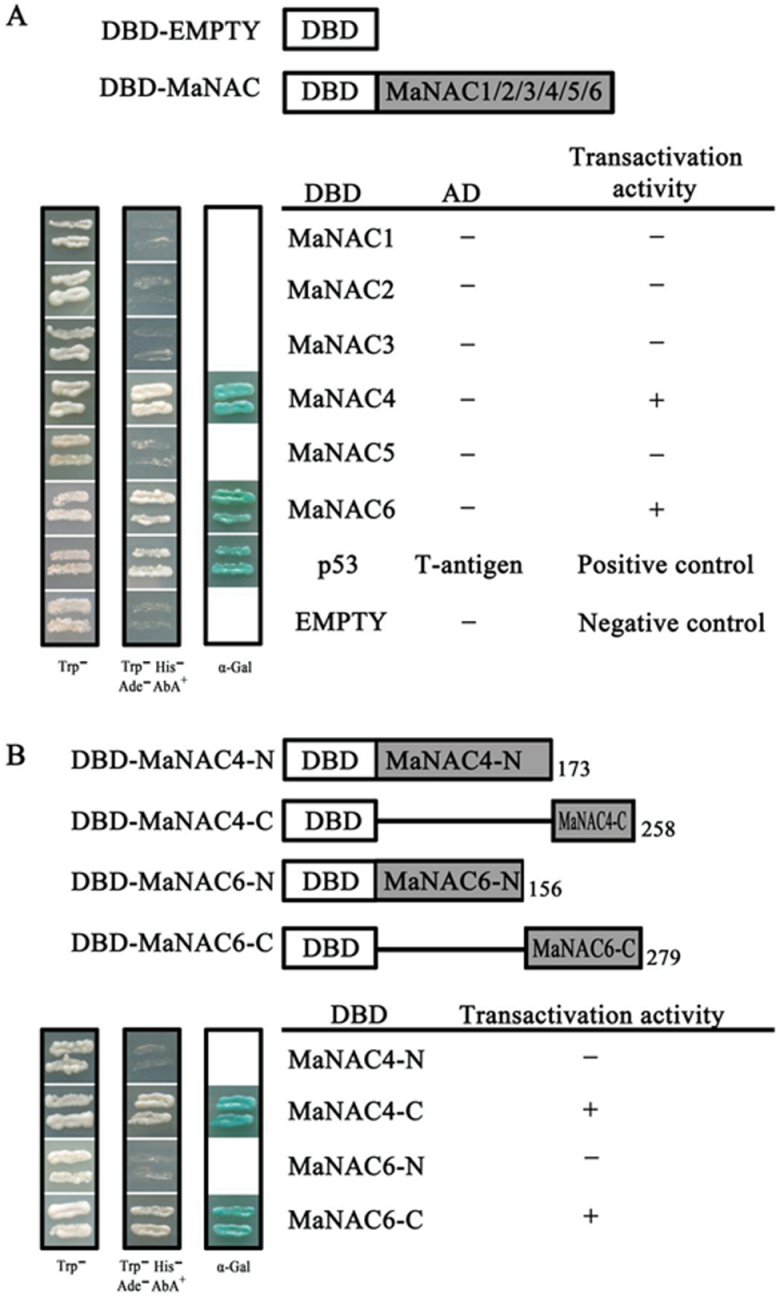
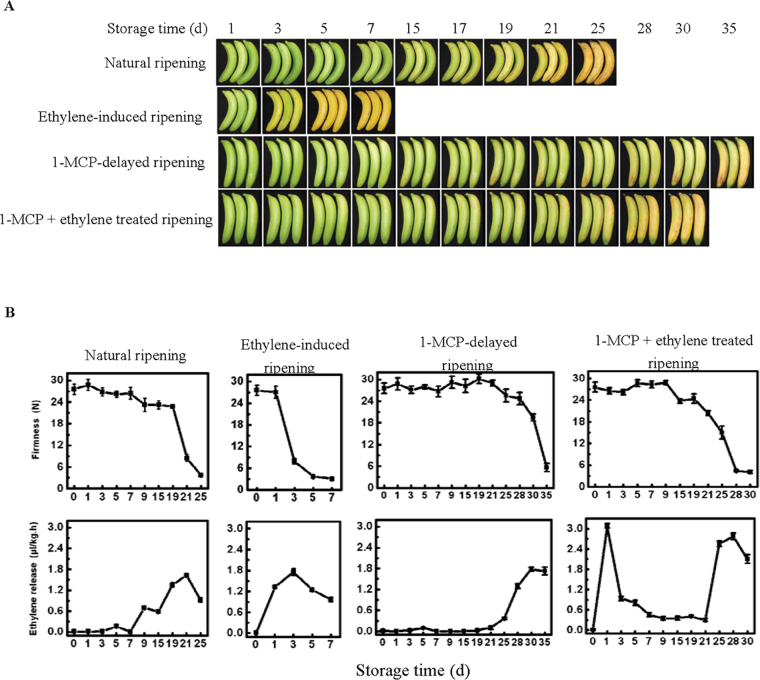
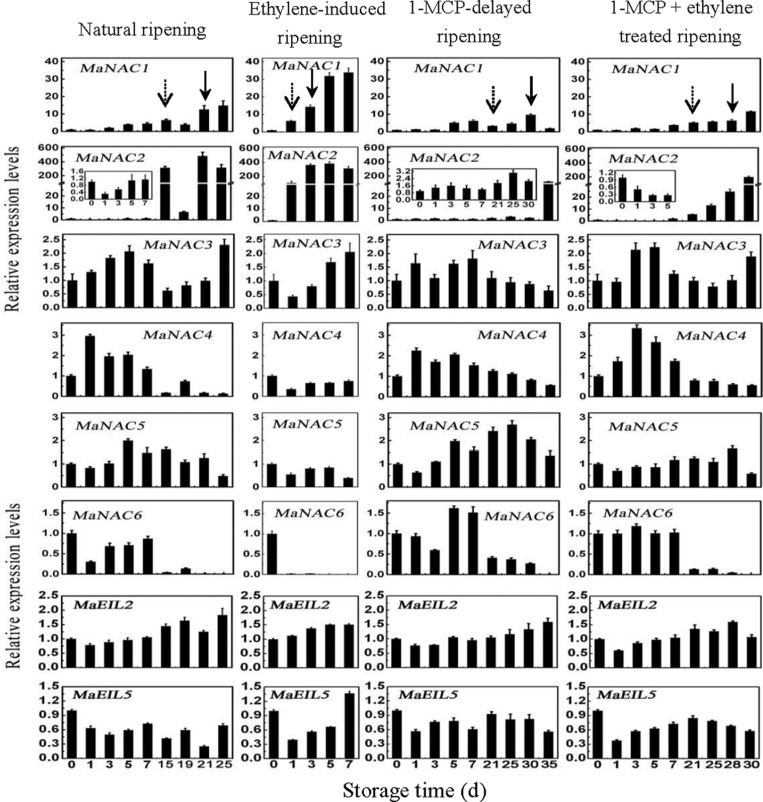
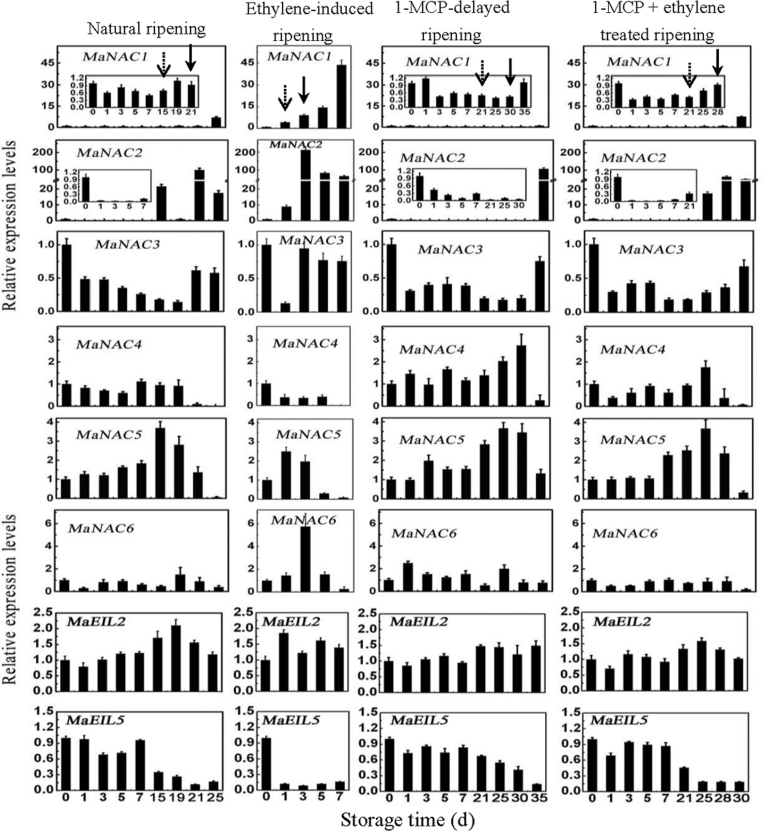
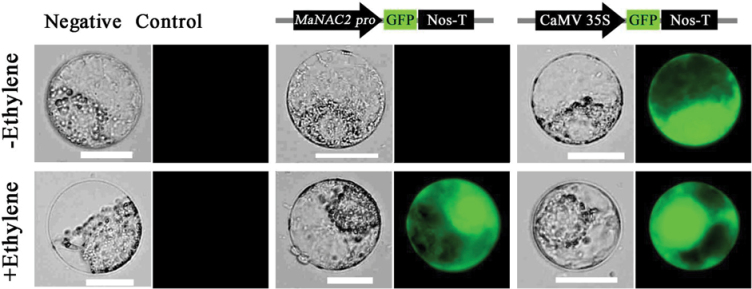
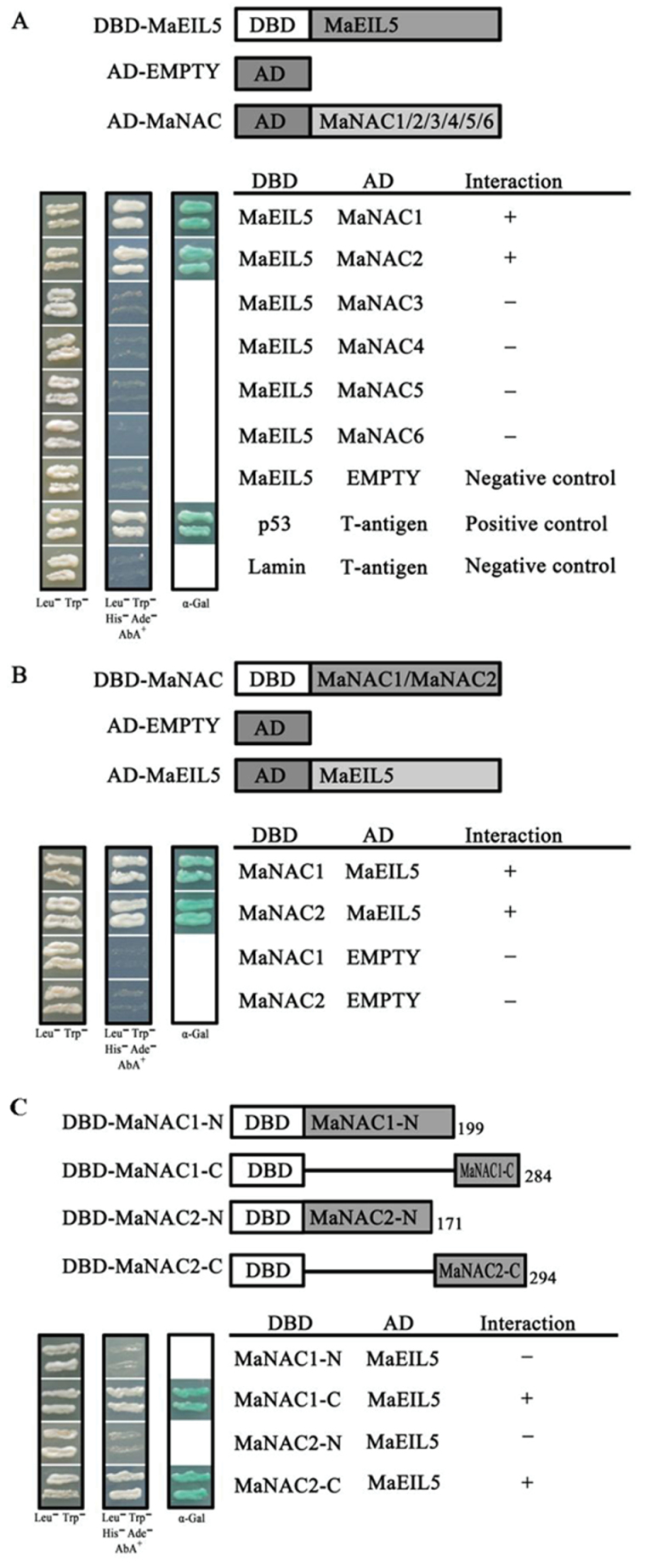
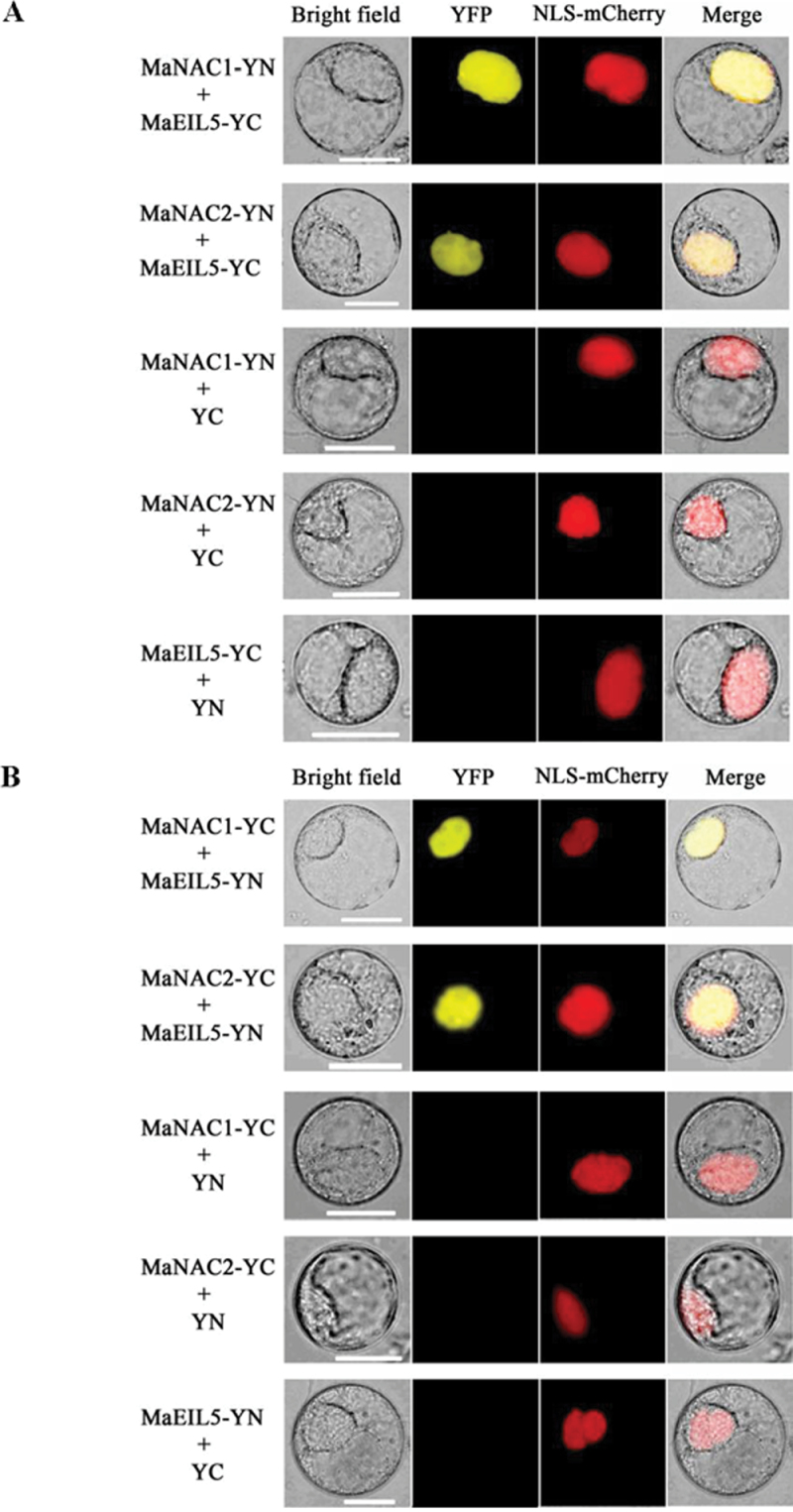
The plant-specific NAC (NAM, ATAF1/2, and CUC2) transcription factors (TFs) play important roles in plant growth, development, and stress responses. However, the precise role of NAC TFs in relation to fruit ripening is poorly understood. In this study, six NAC genes, designated MaNAC1-MaNAC6, were isolated and characterized from banana fruit. Subcellular localization showed that MaNAC1-MaNAC5 proteins localized preferentially to the nucleus, while MaNAC6 was distributed throughout the entire cell. A transactivation assay in yeast demonstrated that MaNAC4 and MaNAC6, as well as their C-terminal regions, possessed trans-activation activity. Gene expression profiles in fruit with four different ripening characteristics, including natural, ethylene-induced, 1-methylcyclopropene (1-MCP)-delayed, and a combination of 1-MCP with ethylene treatment, revealed that the MaNAC genes were differentially expressed in peel and pulp during post-harvest ripening. MaNAC1 and MaNAC2 were apparently upregulated by ethylene in peel and pulp, consistent with the increase in ethylene production. In contrast, MaNAC3 in peel and pulp and MaNAC5 in peel were constitutively expressed, and transcripts of MaNAC4 in peel and pulp and MaNAC6 in peel decreased, while MaNAC5 or MaNAC6 in pulp increased slightly during fruit ripening. Furthermore, the MaNAC2 promoter was activated after ethylene application, further enhancing the involvement of MaNAC2 in fruit ripening. More importantly, yeast two-hybrid and bimolecular fluorescence complementation analyses confirmed that MaNAC1/2 physically interacted with a downstream component of ethylene signalling, ethylene insensitive 3 (EIN3)-like protein, termed MaEIL5, which was downregulated during ripening. Taken together, these results suggest that MaNACs such as MaNAC1/MaNAC2, may be involved in banana fruit ripening via interaction with ethylene signalling components.
Figures

References
-
- Abel S, Theologis A. 1994. Transient transformation of Arabidopsis leaf protoplasts: a versatile experimental system to study gene expression The Plant Journal 5 421–427 - PubMed
-
- Al-Daoud F, Cameron RK. 2011. ANAC055 and ANAC092 contribute non-redundantly in an EIN2-dependent manner to age-related resistance in Arabidopsis Physiological and Molecular Plant Pathology 76 212–222
-
- Bapat VA, Trivedi PK, Ghosh A, Sane VA. 2010. Ripening of fleshy fruit: molecular insight and the role of ethylene Biotechnology Advances 28 94–107 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
