

Clinical, virological and biochemical evidence supporting the association of HIV-1 reverse transcriptase polymorphism R284K and thymidine analogue resistance mutations M41L, L210W and T215Y in patients failing tenofovir/emtricitabine therapy
- PMID: 22889300
- PMCID: PMC3468358
- DOI: 10.1186/1742-4690-9-68
Clinical, virological and biochemical evidence supporting the association of HIV-1 reverse transcriptase polymorphism R284K and thymidine analogue resistance mutations M41L, L210W and T215Y in patients failing tenofovir/emtricitabine therapy
Abstract
Background: Thymidine analogue resistance mutations (TAMs) selected under treatment with nucleoside analogues generate two distinct genotypic profiles in the HIV-1 reverse transcriptase (RT): (i) TAM1: M41L, L210W and T215Y, and (ii) TAM2: D67N, K70R and K219E/Q, and sometimes T215F. Secondary mutations, including thumb subdomain polymorphisms (e.g. R284K) have been identified in association with TAMs. We have identified mutational clusters associated with virological failure during salvage therapy with tenofovir/emtricitabine-based regimens. In this context, we have studied the role of R284K as a secondary mutation associated with mutations of the TAM1 complex.
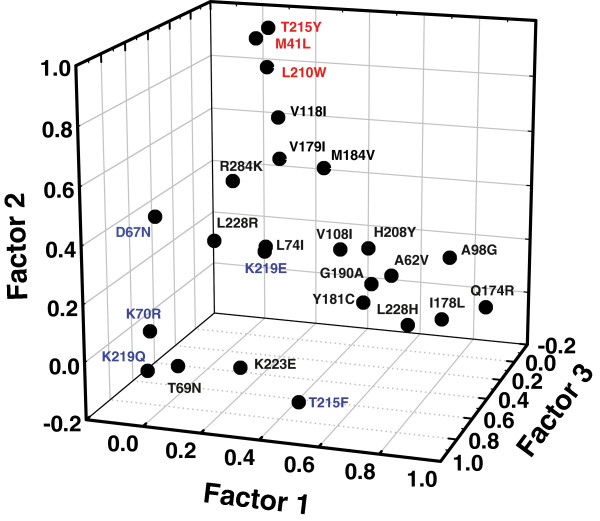
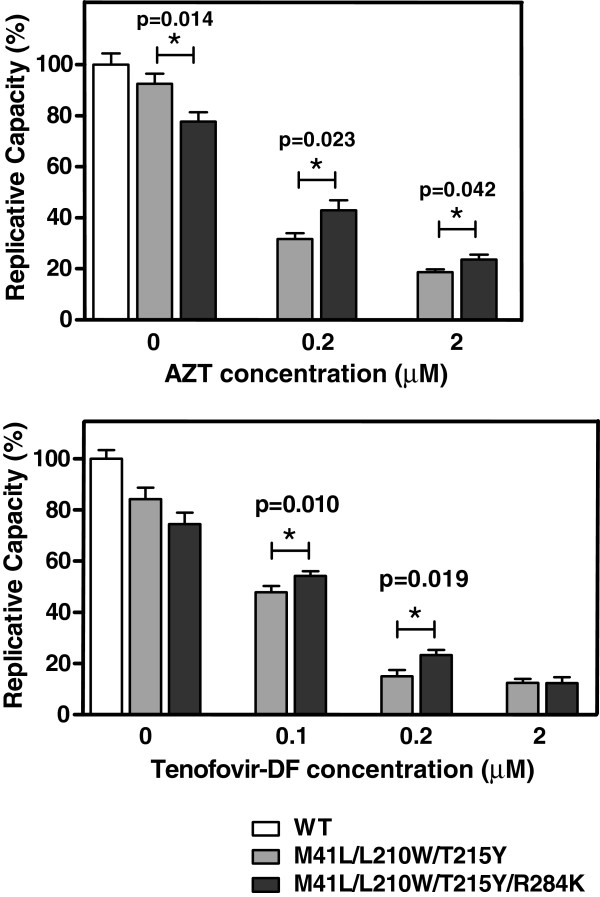
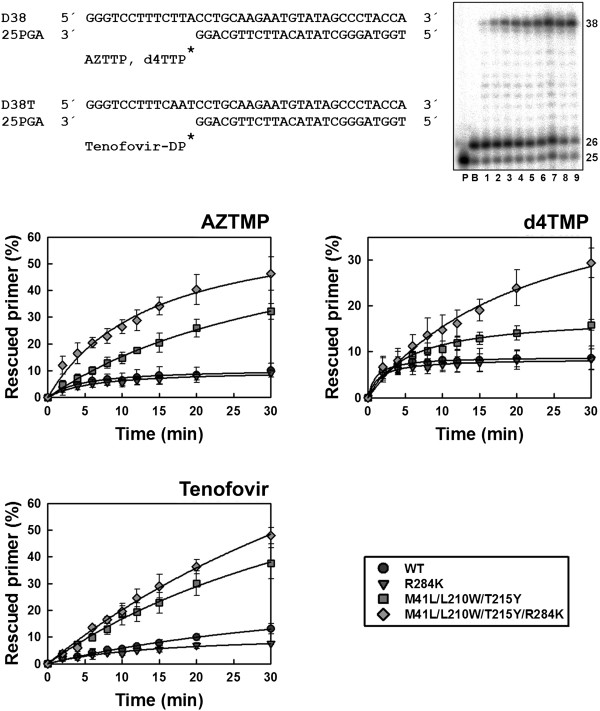
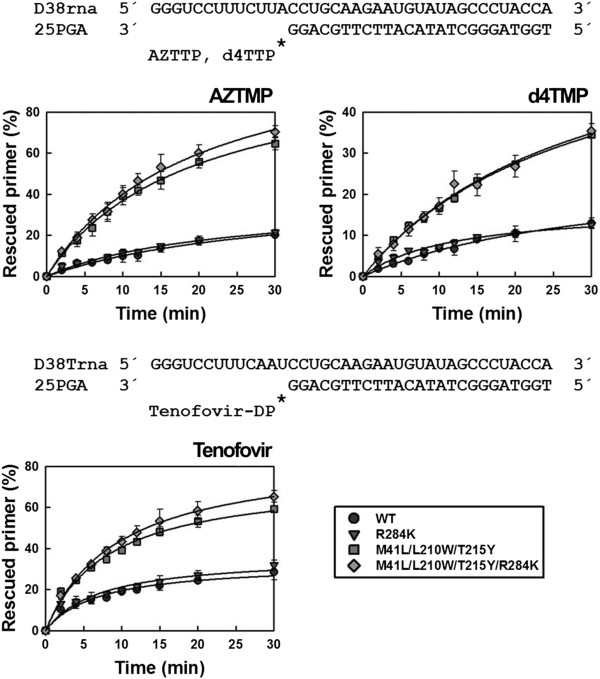
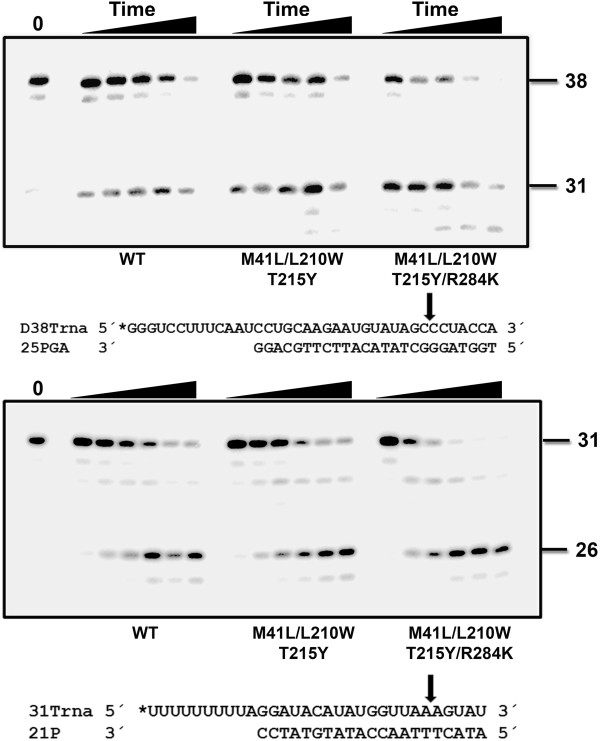
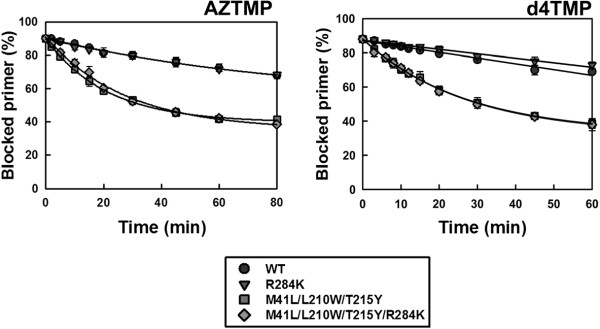
Results: The cross-sectional study carried out with > 200 HIV-1 genotypes showed that virological failure to tenofovir/emtricitabine was strongly associated with the presence of M184V (P < 10-10) and TAMs (P < 10-3), while K65R was relatively uncommon in previously-treated patients failing antiretroviral therapy. Clusters of mutations were identified, and among them, the TAM1 complex showed the highest correlation coefficients. Covariation of TAM1 mutations and V118I, V179I, M184V and R284K was observed. Virological studies showed that the combination of R284K with TAM1 mutations confers a fitness advantage in the presence of zidovudine or tenofovir. Studies with recombinant HIV-1 RTs showed that when associated with TAM1 mutations, R284K had a minimal impact on zidovudine or tenofovir inhibition, and in their ability to excise the inhibitors from blocked DNA primers. However, the mutant RT M41L/L210W/T215Y/R284K showed an increased catalytic rate for nucleotide incorporation and a higher RNase H activity in comparison with WT and mutant M41L/L210W/T215Y RTs. These effects were consistent with its enhanced chain-terminated primer rescue on DNA/DNA template-primers, but not on RNA/DNA complexes, and can explain the higher fitness of HIV-1 having TAM1/R284K mutations.
Conclusions: Our study shows the association of R284K and TAM1 mutations in individuals failing therapy with tenofovir/emtricitabine, and unveils a novel mechanism by which secondary mutations are selected in the context of drug-resistance mutations.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
