

Characterizing circular peptides in mixtures: sequence fragment assembly of cyclotides from a violet plant by MALDI-TOF/TOF mass spectrometry
- PMID: 22890611
- PMCID: PMC3549257
- DOI: 10.1007/s00726-012-1376-x
Characterizing circular peptides in mixtures: sequence fragment assembly of cyclotides from a violet plant by MALDI-TOF/TOF mass spectrometry
Abstract
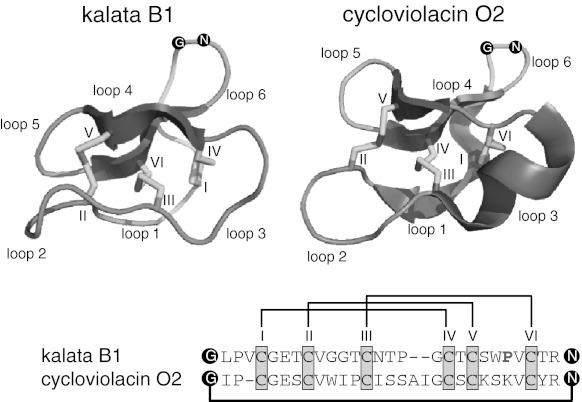
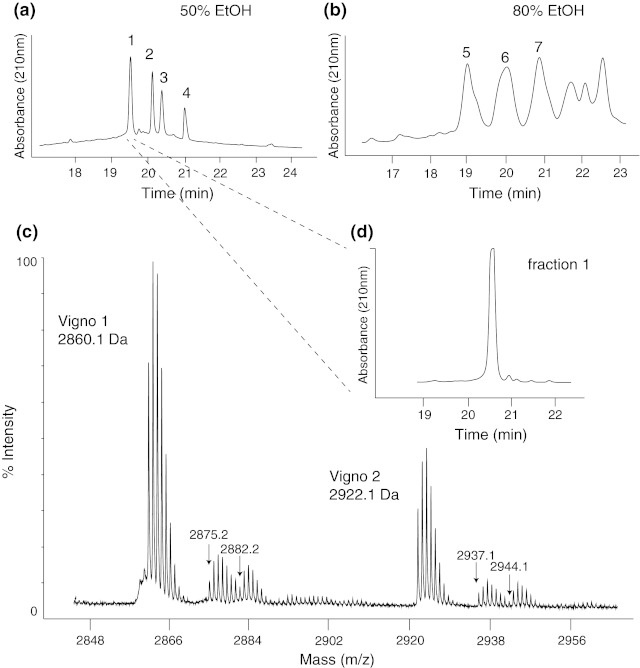
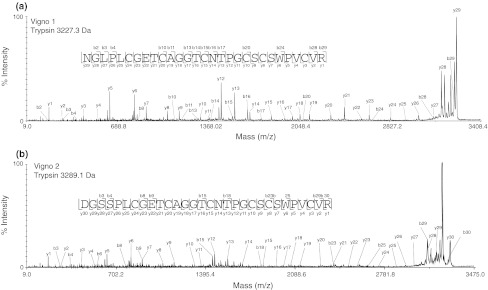
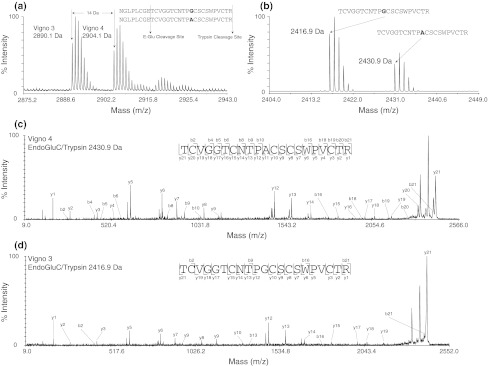
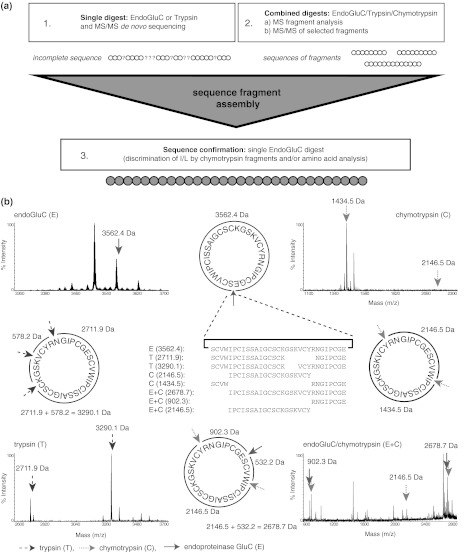
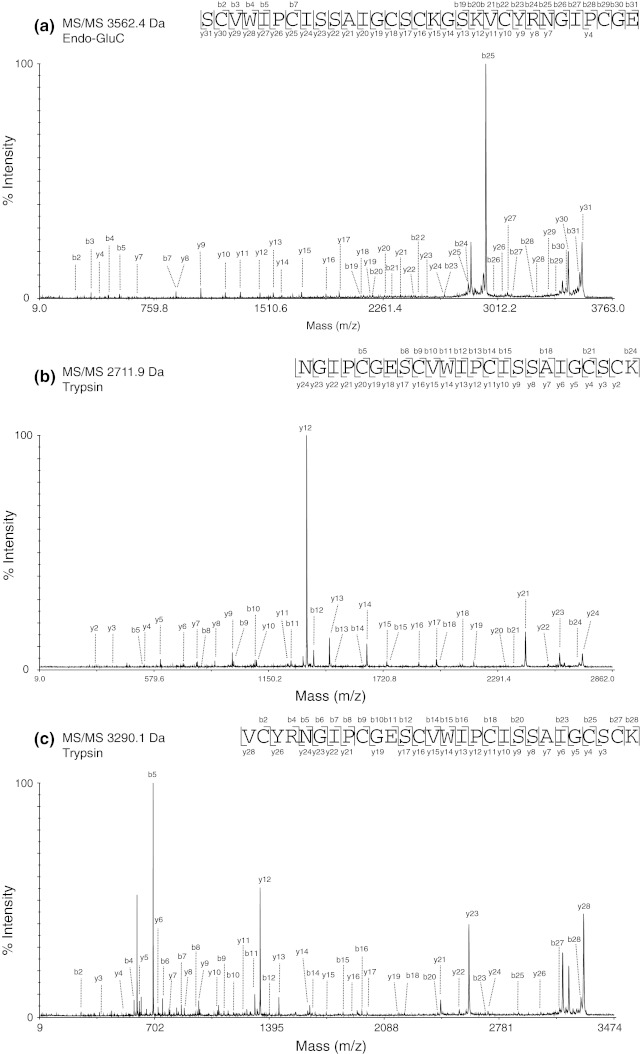
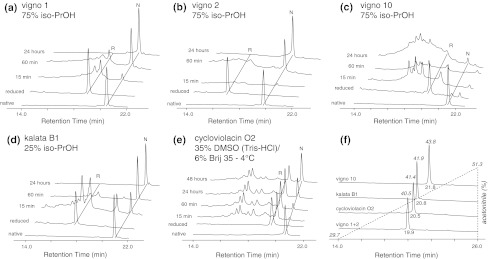
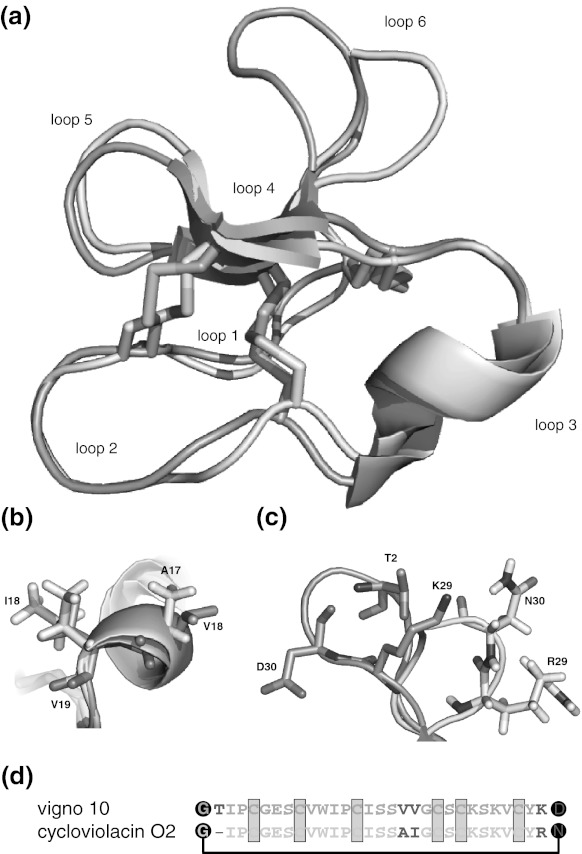
Cyclotides are a very abundant class of plant peptides that display significant sequence variability around a conserved cystine-knot motif and a head-to-tail cyclized backbone conferring them with remarkable stability. Their intrinsic bioactivities combined with tools of peptide engineering make cyclotides an interesting template for the design of novel agrochemicals and pharmaceuticals. However, laborious isolation and purification prior to de novo sequencing limits their discovery and hence their use as scaffolds for peptide-based drug development. Here we extend the knowledge about their sequence diversity by analysing the cyclotide content of a violet species native to Western Asia and the Caucasus region. Using an experimental approach, which was named sequence fragment assembly by MALDI-TOF/TOF, it was possible to characterize 13 cyclotides from Viola ignobilis, whereof ten (vigno 1-10) display previously unknown sequences. Amino acid sequencing of various enzymatic digests of cyclotides allowed the accurate assembly and alignment of smaller fragments to elucidate their primary structure, even when analysing mixtures containing multiple peptides. As a model to further dissect the combinatorial nature of the cyclotide scaffold, we employed in vitro oxidative refolding of representative vigno cyclotides and confirmed the high dependency of folding yield on the inter-cysteine loop sequences. Overall this work highlights the immense structural diversity and plasticity of the unique cyclotide framework. The presented approach for the sequence analysis of peptide mixtures facilitates and accelerates the discovery of novel plant cyclotides.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
