

Exercise- and training-induced upregulation of skeletal muscle fatty acid oxidation are not solely dependent on mitochondrial machinery and biogenesis
- PMID: 22890711
- PMCID: PMC3784190
- DOI: 10.1113/jphysiol.2012.238451
Exercise- and training-induced upregulation of skeletal muscle fatty acid oxidation are not solely dependent on mitochondrial machinery and biogenesis
Abstract
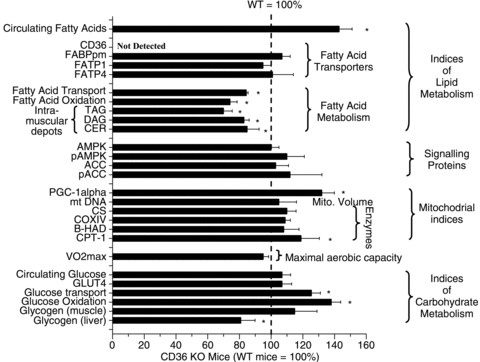
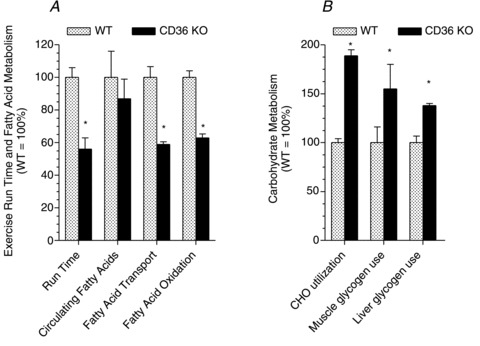
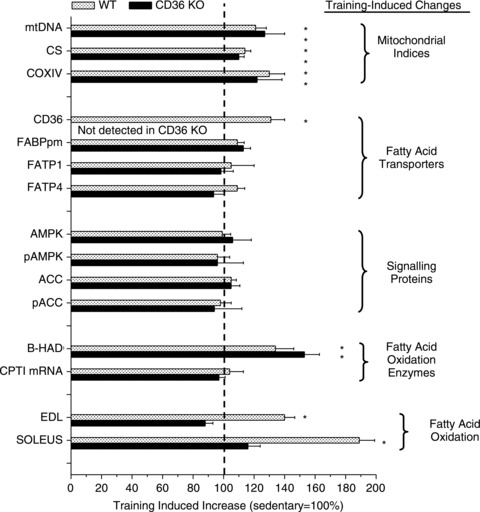
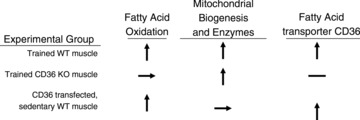
Regulation of skeletal muscle fatty acid oxidation (FAO) and adaptation to exercise training have long been thought to depend on delivery of fatty acids (FAs) to muscle, their diffusion into muscle, and muscle mitochondrial content and biochemical machinery. However, FA entry into muscle occurs via a regulatable, protein-mediated mechanism, involving several transport proteins. Among these CD36 is key. Muscle contraction and pharmacological agents induce CD36 to translocate to the cell surface, a response that regulates FA transport, and hence FAO. In exercising CD36 KO mice, exercise duration (-44%), and FA transport (-41%) and oxidation (-37%) are comparably impaired, while carbohydrate metabolism is augmented. In trained CD36 KO mice, training-induced upregulation of FAO is not observed, despite normal training-induced increases in mitochondrial density and enzymes. Transfecting CD36 into sedentary WT muscle (+41%), comparable to training-induced CD36 increases (+44%) in WT muscle, markedly upregulates FAO to rates observed in trained WT mice, but without any changes in mitochondrial density and enzymes. Evidently, in vivo CD36-mediated FA transport is key for muscle fuel selection and training-induced FAO upregulation, independent of mitochondrial adaptations. This CD36 molecular mechanism challenges the view that skeletal muscle FAO is solely regulated by muscle mitochondrial content and machinery.
Figures

Similar articles
-
In vivo, fatty acid translocase (CD36) critically regulates skeletal muscle fuel selection, exercise performance, and training-induced adaptation of fatty acid oxidation.J Biol Chem. 2012 Jul 6;287(28):23502-16. doi: 10.1074/jbc.M111.315358. Epub 2012 May 14. J Biol Chem. 2012. PMID: 22584574 Free PMC article.
-
Caffeine-stimulated fatty acid oxidation is blunted in CD36 null mice.Acta Physiol (Oxf). 2012 May;205(1):71-81. doi: 10.1111/j.1748-1716.2012.02396.x. Acta Physiol (Oxf). 2012. PMID: 22463611
-
FAT/CD36-null mice reveal that mitochondrial FAT/CD36 is required to upregulate mitochondrial fatty acid oxidation in contracting muscle.Am J Physiol Regul Integr Comp Physiol. 2009 Oct;297(4):R960-7. doi: 10.1152/ajpregu.91021.2008. Epub 2009 Jul 22. Am J Physiol Regul Integr Comp Physiol. 2009. PMID: 19625692
-
Regulation of skeletal muscle mitochondrial fatty acid metabolism in lean and obese individuals.Am J Clin Nutr. 2009 Jan;89(1):455S-62S. doi: 10.3945/ajcn.2008.26717B. Epub 2008 Dec 3. Am J Clin Nutr. 2009. PMID: 19056573 Review.
-
Contribution of FAT/CD36 to the regulation of skeletal muscle fatty acid oxidation: an overview.Acta Physiol (Oxf). 2008 Dec;194(4):293-309. doi: 10.1111/j.1748-1716.2008.01878.x. Epub 2008 Jul 9. Acta Physiol (Oxf). 2008. PMID: 18510711 Review.
Cited by
-
Ca2+ Binding/Permeation via Calcium Channel, CaV1.1, Regulates the Intracellular Distribution of the Fatty Acid Transport Protein, CD36, and Fatty Acid Metabolism.J Biol Chem. 2015 Sep 25;290(39):23751-65. doi: 10.1074/jbc.M115.643544. Epub 2015 Aug 5. J Biol Chem. 2015. PMID: 26245899 Free PMC article.
-
Interplay between Cultured Human Osteoblastic and Skeletal Muscle Cells: Effects of Conditioned Media on Glucose and Fatty Acid Metabolism.Biomedicines. 2023 Oct 27;11(11):2908. doi: 10.3390/biomedicines11112908. Biomedicines. 2023. PMID: 38001909 Free PMC article.
-
Changes in Fat Oxidation and Body Composition after Combined Exercise Intervention in Sedentary Obese Chinese Adults.J Clin Med. 2022 Feb 18;11(4):1086. doi: 10.3390/jcm11041086. J Clin Med. 2022. PMID: 35207356 Free PMC article.
-
CD36 deletion ameliorates diabetic kidney disease by restoring fatty acid oxidation and improving mitochondrial function.Ren Fail. 2023;45(2):2292753. doi: 10.1080/0886022X.2023.2292753. Epub 2023 Dec 14. Ren Fail. 2023. PMID: 38097943 Free PMC article.
-
Reduced efficiency of sarcolipin-dependent respiration in myocytes from humans with severe obesity.Obesity (Silver Spring). 2015 Jul;23(7):1440-9. doi: 10.1002/oby.21123. Epub 2015 May 13. Obesity (Silver Spring). 2015. PMID: 25970801 Free PMC article.
References
-
- Abumrad NA, El-Maghrabi MR, Amri E-Z, Lopez E, Grimaldi P. Cloning of a rat adipocyte membrane protein implicated in binding or transport of long chain fatty acids that is induced during preadipocyte differentiation. Homology with human CD36. J Biol Chem. 1993;268:17665–17668. - PubMed
-
- Aoi W, Naito Y, Takanami Y, Ishii T, Kawai Y, Akagiri S, Kato Y, Osawa T, Yoshikawa T. Astaxanthin improves muscle lipid metabolism in exercise via inhibitory effect of oxidative CPT I modification. Biochem Biophys Res Commun. 2008;366:892–897. - PubMed
-
- Benton CR, Holloway GP, Han XX, Yoshida Y, Snook LA, Lally J, Glatz JF, Luiken JJ, Chabowski A, Bonen A. Increased levels of peroxisome proliferator-activated receptor gamma, coactivator 1 alpha (PGC-1α) improve lipid utilisation, insulin signalling and glucose transport in skeletal muscle of lean and insulin-resistant obese Zucker rats. Diabetologia. 2010;53:2008–2019. - PubMed
-
- Benton CR, Nickerson J, Lally J, Han X-X, Holloway GP, Glatz JFC, Luiken JJFP, Graham TE, Heikkila JJ, Bonen A. Modest PGC-1α overexpression in muscle in vivo is sufficient to increase insulin sensitivity and palmitate oxidation in SS, not IMF, mitochondria. J Biol Chem. 2008;283:4228–4240. - PubMed
-
- Bergstrom J, Hermansen L, Hultman E, Saltin B. Diet, muscle glycogen and physical performance. Acta Physiol Scand. 1967;71:140–150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous