

Disruption of the selenocysteine lyase-mediated selenium recycling pathway leads to metabolic syndrome in mice
- PMID: 22890841
- PMCID: PMC3457337
- DOI: 10.1128/MCB.00293-12
Disruption of the selenocysteine lyase-mediated selenium recycling pathway leads to metabolic syndrome in mice
Abstract
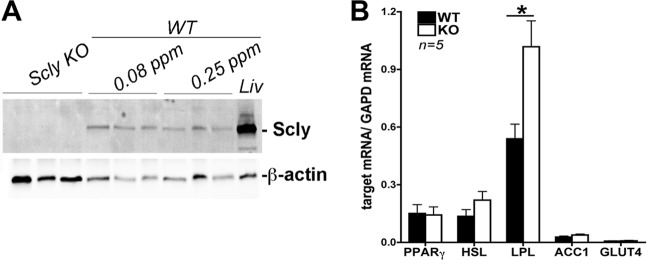
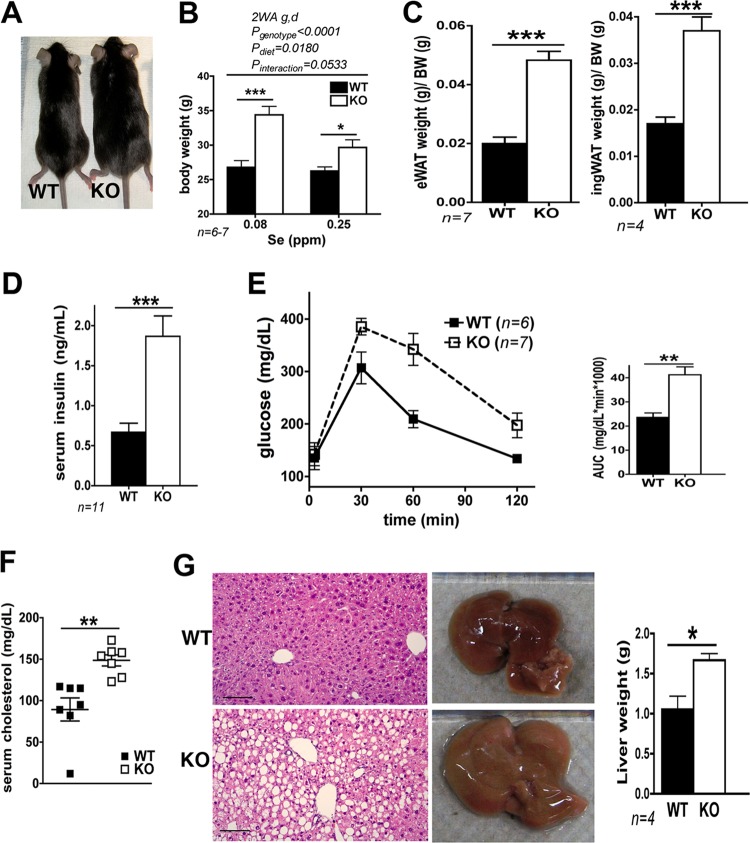
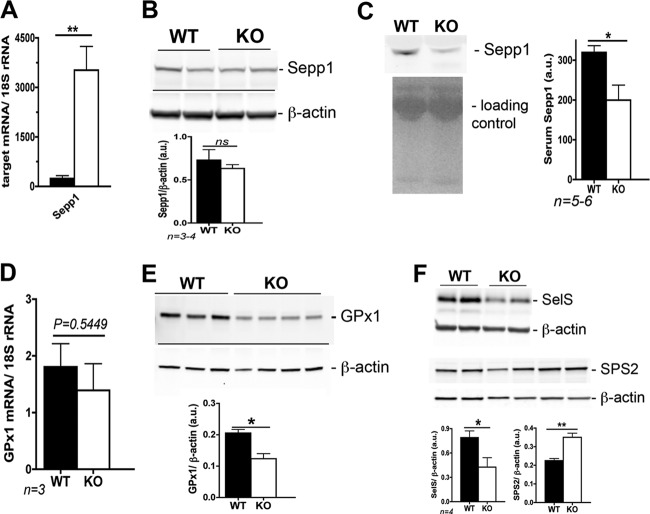
Selenium (Se) is an essential trace element used for biosynthesis of selenoproteins and is acquired either through diet or cellular recycling mechanisms. Selenocysteine lyase (Scly) is the enzyme that supplies Se for selenoprotein biosynthesis via decomposition of the amino acid selenocysteine (Sec). Knockout (KO) of Scly in a mouse affected hepatic glucose and lipid homeostasis. Mice lacking Scly and raised on an Se-adequate diet exhibit hyperinsulinemia, hyperleptinemia, glucose intolerance, and hepatic steatosis, with increased hepatic oxidative stress, but maintain selenoprotein levels and circulating Se status. Insulin challenge of Scly KO mice results in attenuated Akt phosphorylation but does not decrease phosphorylation levels of AMP kinase alpha (AMPKα). Upon dietary Se restriction, Scly KO animals develop several characteristics of metabolic syndrome, such as obesity, fatty liver, and hypercholesterolemia, with aggravated hyperleptinemia, hyperinsulinemia, and glucose intolerance. Hepatic glutathione peroxidase 1 (GPx1) and selenoprotein S (SelS) production and circulating selenoprotein P (Sepp1) levels are significantly diminished. Scly disruption increases the levels of insulin-signaling inhibitor PTP1B. Our results suggest a dependence of glucose and lipid homeostasis on Scly activity. These findings connect Se and energy metabolism and demonstrate for the first time a unique physiological role of Scly in an animal model.
Figures







Similar articles
-
Diet-induced obesity in the selenocysteine lyase knockout mouse.Antioxid Redox Signal. 2015 Oct 1;23(10):761-74. doi: 10.1089/ars.2015.6277. Epub 2015 Aug 24. Antioxid Redox Signal. 2015. PMID: 26192035 Free PMC article.
-
Effects of selenium supplementation on diet-induced obesity in mice with a disruption of the selenocysteine lyase gene.J Trace Elem Med Biol. 2020 Dec;62:126596. doi: 10.1016/j.jtemb.2020.126596. Epub 2020 Jul 11. J Trace Elem Med Biol. 2020. PMID: 32683228 Free PMC article.
-
Interference of selenium and selenoproteins with the insulin-regulated carbohydrate and lipid metabolism.Free Radic Biol Med. 2013 Dec;65:1538-1547. doi: 10.1016/j.freeradbiomed.2013.07.016. Epub 2013 Jul 18. Free Radic Biol Med. 2013. PMID: 23872396 Review.
-
Combined Omics Reveals That Disruption of the Selenocysteine Lyase Gene Affects Amino Acid Pathways in Mice.Nutrients. 2019 Oct 26;11(11):2584. doi: 10.3390/nu11112584. Nutrients. 2019. PMID: 31717805 Free PMC article.
-
Selenium and diabetes--evidence from animal studies.Free Radic Biol Med. 2013 Dec;65:1548-1556. doi: 10.1016/j.freeradbiomed.2013.07.012. Epub 2013 Jul 16. Free Radic Biol Med. 2013. PMID: 23867154 Free PMC article. Review.
Cited by
-
Selenium Level and Dyslipidemia in Rural Elderly Chinese.PLoS One. 2015 Sep 18;10(9):e0136706. doi: 10.1371/journal.pone.0136706. eCollection 2015. PLoS One. 2015. PMID: 26380972 Free PMC article.
-
Hypothalamic redox balance and leptin signaling - Emerging role of selenoproteins.Free Radic Biol Med. 2018 Nov 1;127:172-181. doi: 10.1016/j.freeradbiomed.2018.02.038. Epub 2018 Mar 5. Free Radic Biol Med. 2018. PMID: 29518483 Free PMC article. Review.
-
Dietary Selenium Deficiency Partially Mimics the Metabolic Effects of Arsenic.Nutrients. 2021 Aug 23;13(8):2894. doi: 10.3390/nu13082894. Nutrients. 2021. PMID: 34445052 Free PMC article.
-
Selenium status and type 2 diabetes risk.Arch Biochem Biophys. 2022 Nov 15;730:109400. doi: 10.1016/j.abb.2022.109400. Epub 2022 Sep 16. Arch Biochem Biophys. 2022. PMID: 36122760 Free PMC article. Review.
-
Female Mice with Selenocysteine tRNA Deletion in Agrp Neurons Maintain Leptin Sensitivity and Resist Weight Gain While on a High-Fat Diet.Int J Mol Sci. 2021 Oct 12;22(20):11010. doi: 10.3390/ijms222011010. Int J Mol Sci. 2021. PMID: 34681674 Free PMC article.
References
-
- Bjorbaek C, El-Haschimi K, Frantz JD, Flier JS. 1999. The role of SOCS-3 in leptin signaling and leptin resistance. J. Biol. Chem. 274:30059–30065 - PubMed
-
- Boitani C, Puglisi R. 2008. Selenium, a key element in spermatogenesis and male fertility. Adv. Exp. Med. Biol. 636:65–73 - PubMed
-
- Burk RF, Hill KE. 2005. Selenoprotein P: an extracellular protein with unique physical characteristics and a role in selenium homeostasis. Annu. Rev. Nutr. 25:215–235 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous