

Primate-specific melanoma antigen-A11 regulates isoform-specific human progesterone receptor-B transactivation
- PMID: 22891251
- PMCID: PMC3464583
- DOI: 10.1074/jbc.M112.372797
Primate-specific melanoma antigen-A11 regulates isoform-specific human progesterone receptor-B transactivation
Abstract
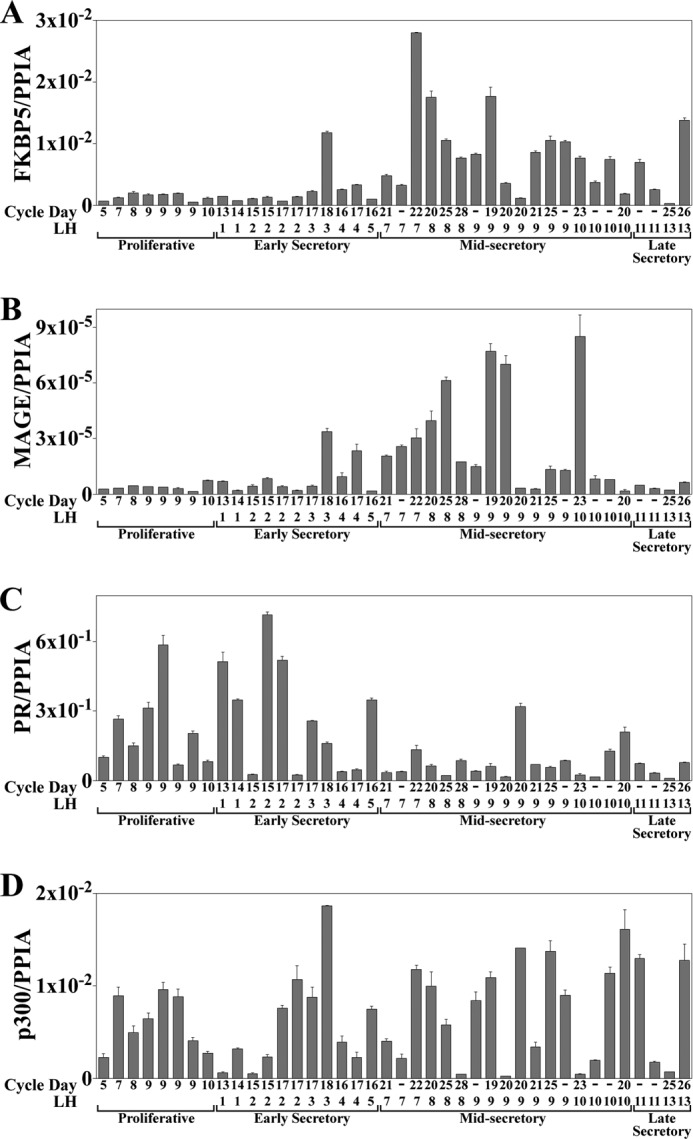
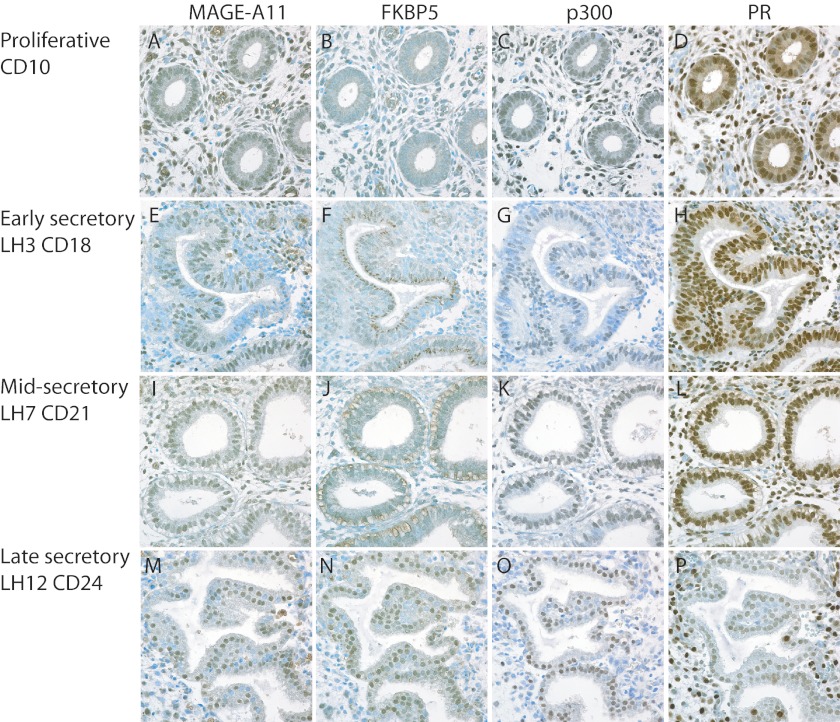
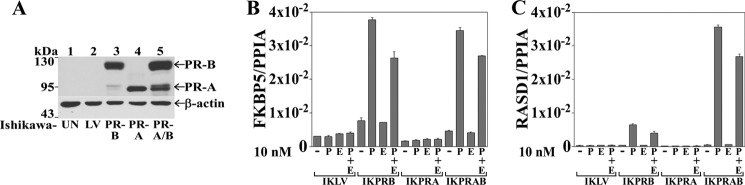
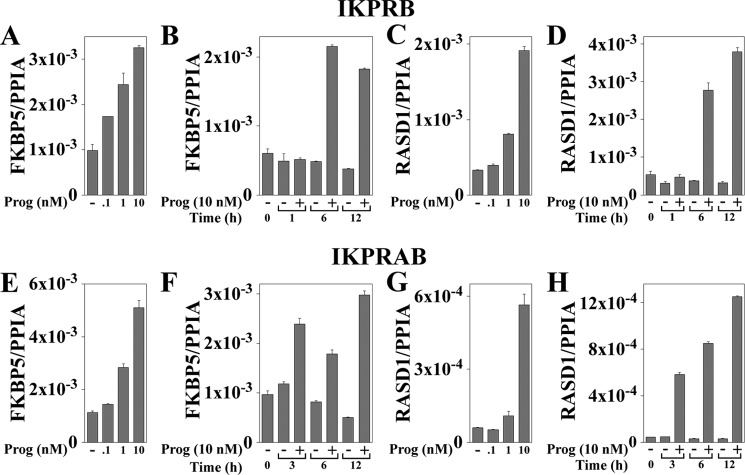
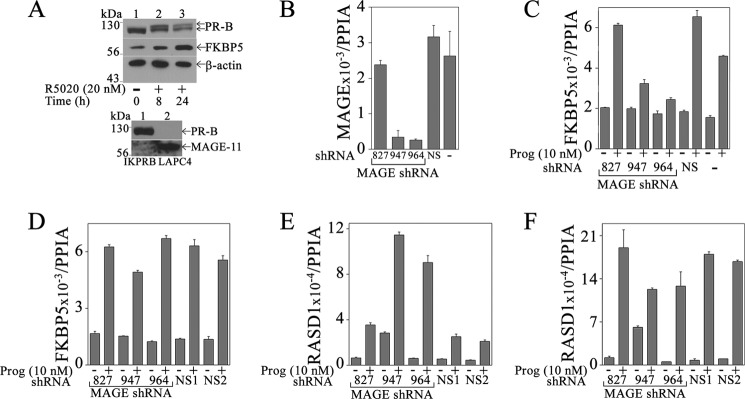
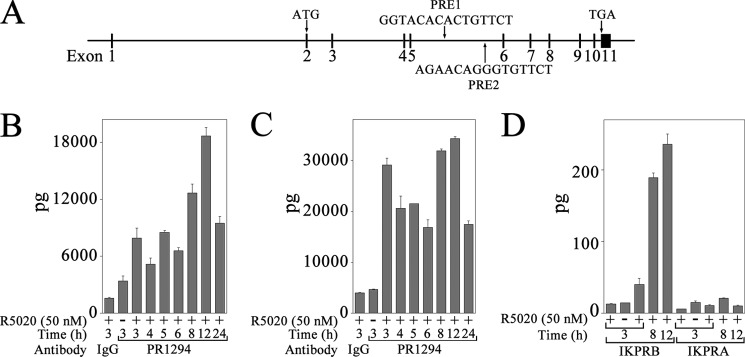
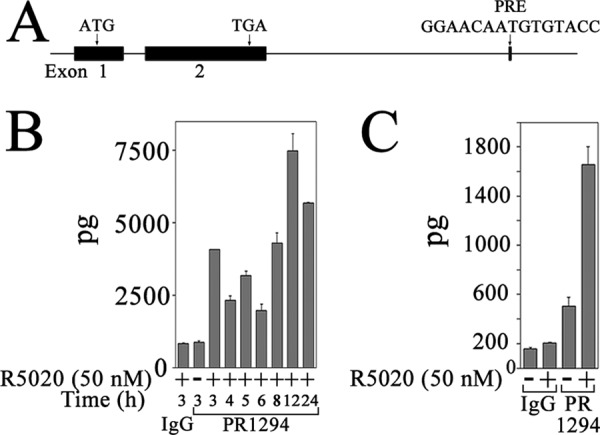
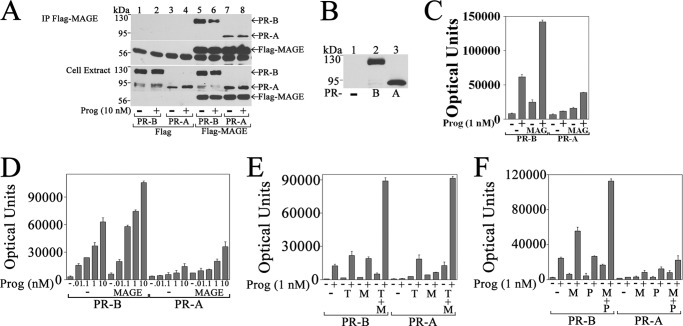
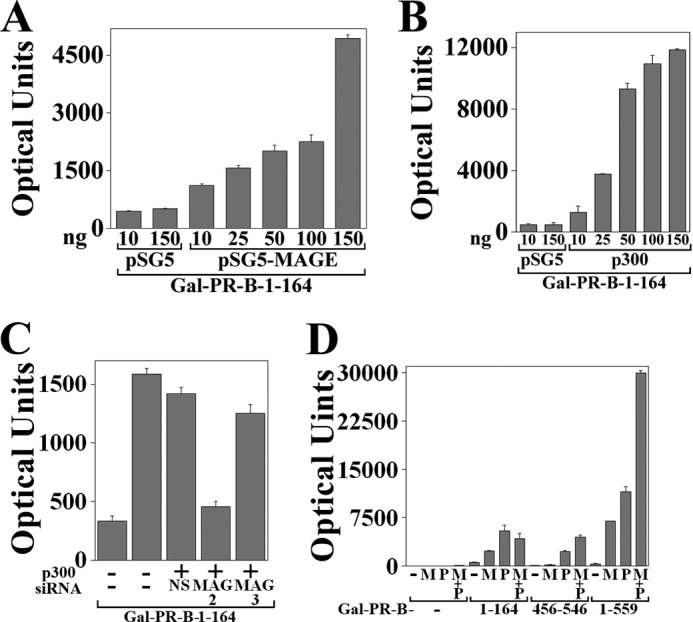
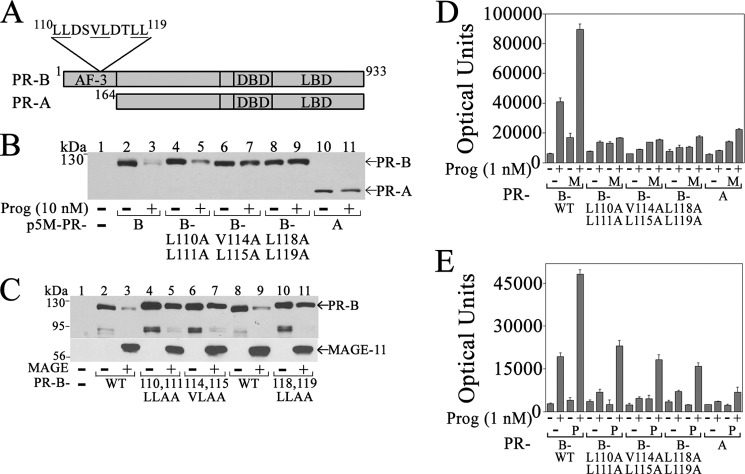
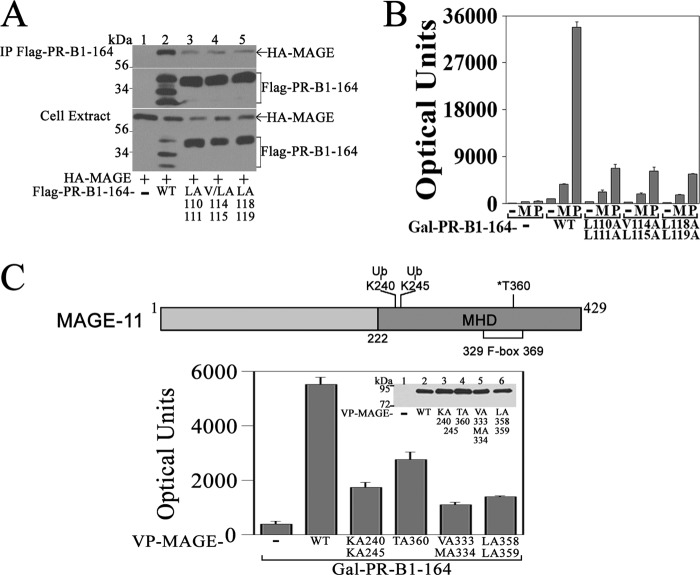
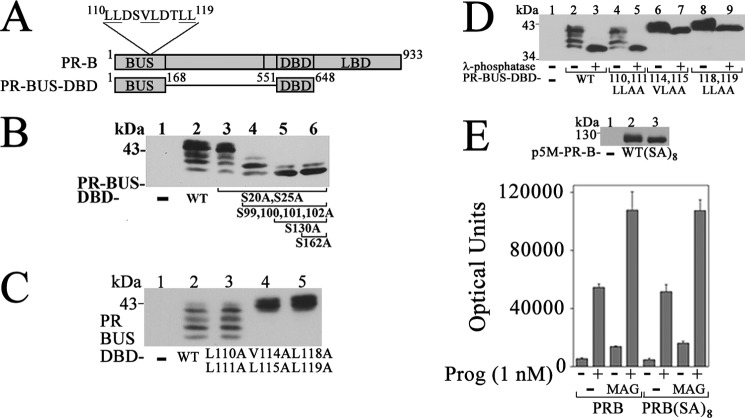
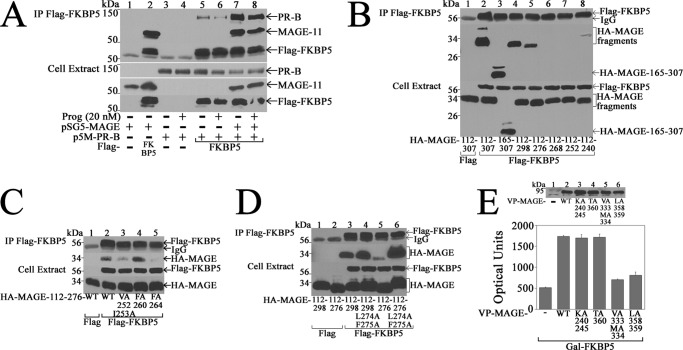
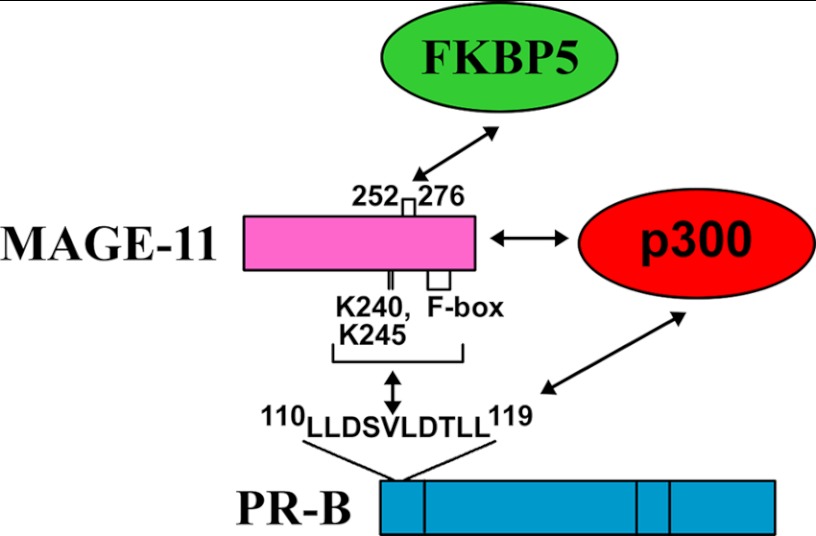
Progesterone acting through the progesterone receptor (PR) and its coregulators prepares the human endometrium for receptivity to embryo implantation and maintains pregnancy. The menstrual cycle-dependent expression of melanoma antigen-A11 (MAGE-11) in the mid-secretory human endometrium suggested a novel function in human PR signaling. Here we show that MAGE-11 is an isoform-specific coregulator responsible for the greater transcriptional activity of human PR-B relative to PR-A. PR was recruited to progesterone response regions of progesterone-regulated FK506-binding protein 5 (FKBP5) immunophilin and small Ras family G protein cell growth inhibitor RASD1 genes. Expression of MAGE-11 lentivirus shRNA in human endometrial Ishikawa cells expressing PR-B showed that MAGE-11 is required for isoform-specific PR-B up-regulation of FKBP5. In contrast, MAGE-11 was not required for progesterone up-regulation of RASD1 in endometrial cells expressing the PR-A/B heterodimer. Target gene specificity of PR-B depended on the synergistic actions of MAGE-11 and p300 mediated by the unique PR-B NH(2)-terminal (110)LLXXVLXXLL(119) motif that interacts with the MAGE-11 F-box region in a phosphorylation- and ubiquitinylation-dependent manner. A progesterone-dependent mechanism is proposed in which MAGE-11 and p300 increase PR-B up-regulation of the FKBP5 gene. MAGE-11 down-regulates PR-B, similar to the effects of progesterone, and interacts with FKBP5 to stabilize a complex with PR-B. We conclude that the coregulator function of MAGE-11 extends to isoform-specific regulation of PR-B during the cyclic development of the human endometrium.
Figures
Similar articles
-
Hormone control and expression of androgen receptor coregulator MAGE-11 in human endometrium during the window of receptivity to embryo implantation.Mol Hum Reprod. 2008 Feb;14(2):107-16. doi: 10.1093/molehr/gam080. Epub 2007 Nov 29. Mol Hum Reprod. 2008. PMID: 18048459 Free PMC article.
-
Gain in transcriptional activity by primate-specific coevolution of melanoma antigen-A11 and its interaction site in androgen receptor.J Biol Chem. 2011 Aug 26;286(34):29951-63. doi: 10.1074/jbc.M111.244715. Epub 2011 Jul 5. J Biol Chem. 2011. PMID: 21730049 Free PMC article.
-
Melanoma antigen-A11 (MAGE-A11) enhances transcriptional activity by linking androgen receptor dimers.J Biol Chem. 2013 Jan 18;288(3):1939-52. doi: 10.1074/jbc.M112.428409. Epub 2012 Nov 21. J Biol Chem. 2013. PMID: 23172223 Free PMC article.
-
Role of nuclear progesterone receptor isoforms in uterine pathophysiology.Hum Reprod Update. 2015 Mar-Apr;21(2):155-73. doi: 10.1093/humupd/dmu056. Epub 2014 Nov 18. Hum Reprod Update. 2015. PMID: 25406186 Free PMC article. Review.
-
Role of progesterone in nonhuman primate implantation.Semin Reprod Med. 2007 Nov;25(6):418-30. doi: 10.1055/s-2007-991039. Semin Reprod Med. 2007. PMID: 17960526 Review.
Cited by
-
Generation of Progesterone-Responsive Endometrial Stromal Fibroblasts from Human Induced Pluripotent Stem Cells: Role of the WNT/CTNNB1 Pathway.Stem Cell Reports. 2018 Nov 13;11(5):1136-1155. doi: 10.1016/j.stemcr.2018.10.002. Epub 2018 Nov 1. Stem Cell Reports. 2018. PMID: 30392973 Free PMC article.
-
Transcription factor FOXD1 and miRNA-204-5p play a major role in B4GALNT2 downregulation in colon cancer.Sci Rep. 2025 Jan 13;15(1):1821. doi: 10.1038/s41598-025-85450-z. Sci Rep. 2025. PMID: 39805916 Free PMC article.
-
Proto-oncogene activity of melanoma antigen-A11 (MAGE-A11) regulates retinoblastoma-related p107 and E2F1 proteins.J Biol Chem. 2013 Aug 23;288(34):24809-24. doi: 10.1074/jbc.M113.468579. Epub 2013 Jul 12. J Biol Chem. 2013. PMID: 23853093 Free PMC article.
-
Emerging roles of the MAGE protein family in stress response pathways.J Biol Chem. 2020 Nov 20;295(47):16121-16155. doi: 10.1074/jbc.REV120.008029. Epub 2020 Sep 13. J Biol Chem. 2020. PMID: 32921631 Free PMC article. Review.
-
MAGEA11 as a STAD Prognostic Biomarker Associated with Immune Infiltration.Diagnostics (Basel). 2022 Oct 16;12(10):2506. doi: 10.3390/diagnostics12102506. Diagnostics (Basel). 2022. PMID: 36292195 Free PMC article.
References
-
- Carson D. D., Bagchi I., Dey S. K., Enders A. C., Fazleabas A. T., Lessey B. A., Yoshinaga K. (2000) Embryo implantation. Dev. Biol. 223, 217–237 - PubMed
-
- Yin P., Lin Z., Reierstad S., Wu J., Ishikawa H., Marsh E. E., Innes J., Cheng Y., Pearson K., Coon J. S., 5th, Kim J. J., Chakravarti D., Bulun S. E. (2010) Transcription factor KLF11 integrates progesterone receptor signaling and proliferation in uterine leiomyoma cells. Cancer Res. 70, 1722–1730 - PMC - PubMed
-
- Chwalisz K., Perez M. C., Demanno D., Winkel C., Schubert G., Elger W. (2005) Selective progesterone receptor modulator development and use in the treatment of leiomyomata and endometriosis. Endocr. Rev. 26, 423–438 - PubMed
-
- Challis J. R. G., Matthews S. G., Gibb W., Lye S. J. (2000) Endocrine and paracrine regulation of birth at term and preterm. Endocr. Rev. 21, 514–550 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
