

Thyroid hormone triggers the developmental loss of axonal regenerative capacity via thyroid hormone receptor α1 and krüppel-like factor 9 in Purkinje cells
- PMID: 22891348
- PMCID: PMC3435228
- DOI: 10.1073/pnas.1119853109
Thyroid hormone triggers the developmental loss of axonal regenerative capacity via thyroid hormone receptor α1 and krüppel-like factor 9 in Purkinje cells
Abstract
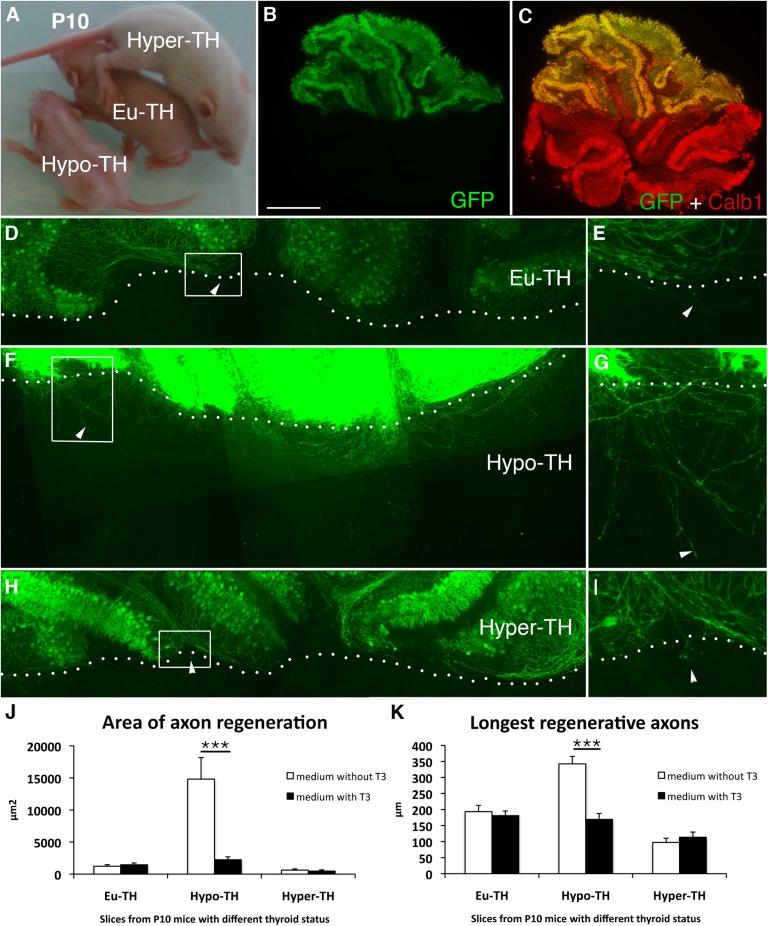
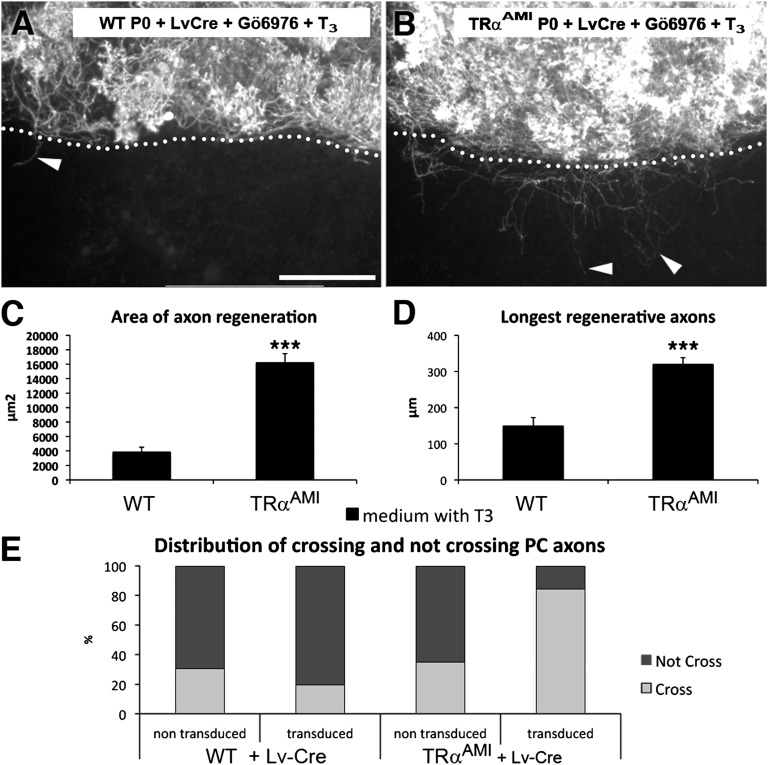
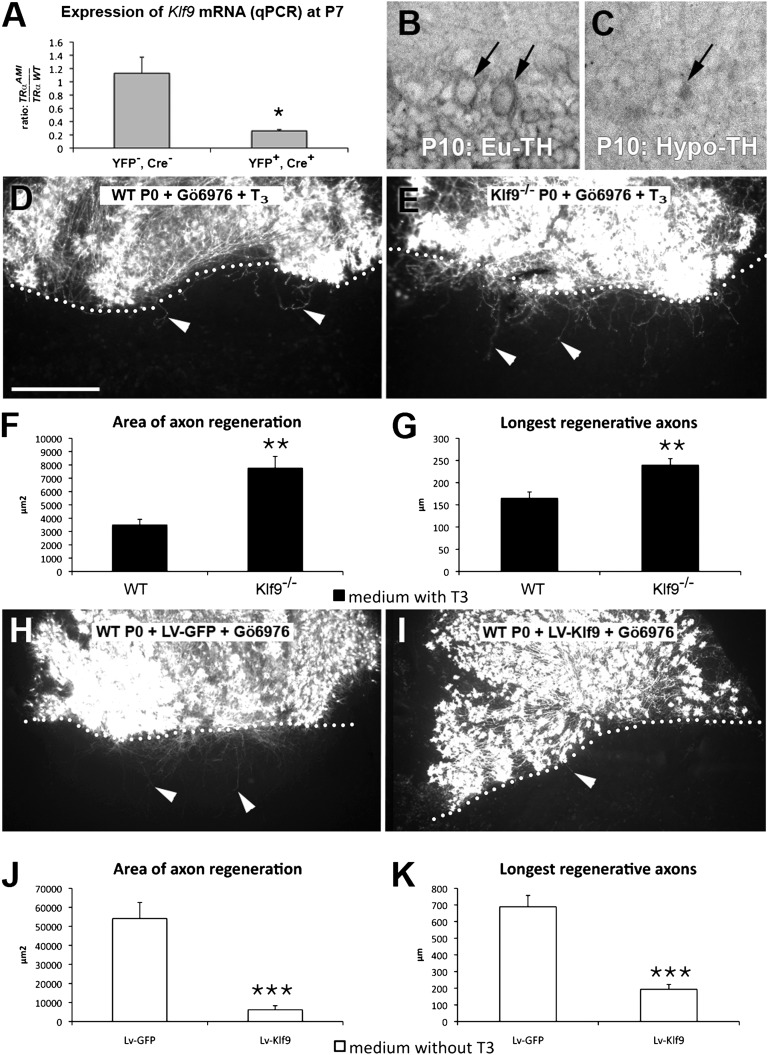
Neurons in the CNS of higher vertebrates lose their ability to regenerate their axons at a stage of development that coincides with peak circulating thyroid hormone (T(3)) levels. Here, we examined whether this peak in T(3) is involved in the loss of axonal regenerative capacity in Purkinje cells (PCs). This event occurs at the end of the first postnatal week in mice. Using organotypic culture, we found that the loss of axon regenerative capacity was triggered prematurely by early exposure of mouse PCs to T(3), whereas it was delayed in the absence of T(3). Analysis of mutant mice showed that this effect was mainly mediated by the T(3) receptor α1. Using gain- and loss-of-function approaches, we also showed that Krüppel-like factor 9 was a key mediator of this effect of T(3). These results indicate that the sudden physiological increase in T(3) during development is involved in the onset of the loss of axon regenerative capacity in PCs. This loss of regenerative capacity might be part of the general program triggered by T(3) throughout the body, which adapts the animal to its postnatal environment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Schwab ME, Bartholdi D. Degeneration and regeneration of axons in the lesioned spinal cord. Physiol Rev. 1996;76:319–370. - PubMed
-
- Schwab ME. Nogo and axon regeneration. Curr Opin Neurobiol. 2004;14(1):118–124. - PubMed
-
- He Z, Koprivica V. The Nogo signaling pathway for regeneration block. Annu Rev Neurosci. 2004;27:341–368. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
