

The Escherichia coli RlmN methyltransferase is a dual-specificity enzyme that modifies both rRNA and tRNA and controls translational accuracy
- PMID: 22891362
- PMCID: PMC3446703
- DOI: 10.1261/rna.033266.112
The Escherichia coli RlmN methyltransferase is a dual-specificity enzyme that modifies both rRNA and tRNA and controls translational accuracy
Abstract
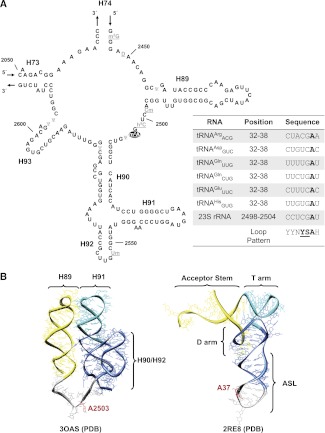
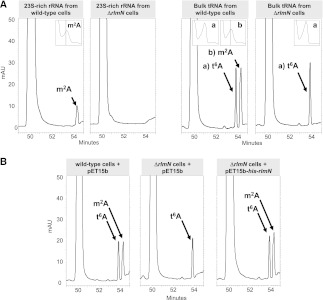
Modifying RNA enzymes are highly specific for substrate-rRNA or tRNA-and the target position. In Escherichia coli, there are very few multisite acting enzymes, and only one rRNA/tRNA dual-specificity enzyme, pseudouridine synthase RluA, has been identified to date. Among the tRNA-modifying enzymes, the methyltransferase responsible for the m(2)A synthesis at purine 37 in a tRNA set still remains unknown. m(2)A is also present at position 2503 in the peptidyl transferase center of 23S RNA, where it is introduced by RlmN, a radical S-adenosyl-L-methionine (SAM) enzyme. Here, we show that E. coli RlmN is a dual-specificity enzyme that catalyzes methylation of both rRNA and tRNA. The ΔrlmN mutant lacks m(2)A in both RNA types, whereas the expression of recombinant RlmN from a plasmid introduced into this mutant restores tRNA modification. Moreover, RlmN performs m(2)A(37) synthesis in vitro using a tRNA chimera as a substrate. This chimera has also proved useful to characterize some tRNA identity determinants for RlmN and other tRNA modification enzymes. Our data suggest that RlmN works in a late step during tRNA maturation by recognizing a precise 3D structure of tRNA. RlmN inactivation increases the misreading of a UAG stop codon. Since loss of m(2)A(37) from tRNA is expected to produce a hyperaccurate phenotype, we believe that the error-prone phenotype exhibited by the ΔrlmN mutant is due to loss of m(2)A from 23S rRNA and, accordingly, that the m(2)A2503 modification plays a crucial role in the proofreading step occurring at the peptidyl transferase center.
Figures


Similar articles
-
Determinants of tRNA Recognition by the Radical SAM Enzyme RlmN.PLoS One. 2016 Nov 30;11(11):e0167298. doi: 10.1371/journal.pone.0167298. eCollection 2016. PLoS One. 2016. PMID: 27902775 Free PMC article.
-
Antibiotic resistance evolved via inactivation of a ribosomal RNA methylating enzyme.Nucleic Acids Res. 2016 Oct 14;44(18):8897-8907. doi: 10.1093/nar/gkw699. Epub 2016 Aug 5. Nucleic Acids Res. 2016. PMID: 27496281 Free PMC article.
-
RlmN and Cfr are radical SAM enzymes involved in methylation of ribosomal RNA.J Am Chem Soc. 2010 Mar 24;132(11):3953-64. doi: 10.1021/ja910850y. J Am Chem Soc. 2010. PMID: 20184321 Free PMC article.
-
Structure and function of ribosomal RNA.Biochem Cell Biol. 1995 Nov-Dec;73(11-12):997-1009. doi: 10.1139/o95-107. Biochem Cell Biol. 1995. PMID: 8722015 Review.
-
What do we know about ribosomal RNA methylation in Escherichia coli?Biochimie. 2015 Oct;117:110-8. doi: 10.1016/j.biochi.2014.11.019. Epub 2014 Dec 13. Biochimie. 2015. PMID: 25511423 Review.
Cited by
-
Radical SAM-mediated methylation reactions.Curr Opin Chem Biol. 2013 Aug;17(4):597-604. doi: 10.1016/j.cbpa.2013.05.032. Epub 2013 Jul 5. Curr Opin Chem Biol. 2013. PMID: 23835516 Free PMC article. Review.
-
Plasticity and conditional essentiality of modification enzymes for domain V of Escherichia coli 23S ribosomal RNA.RNA. 2022 Jun;28(6):796-807. doi: 10.1261/rna.079096.121. Epub 2022 Mar 8. RNA. 2022. PMID: 35260421 Free PMC article.
-
Elp3 and RlmN: A tale of two mitochondrial tail-anchored radical SAM enzymes in Toxoplasma gondii.PLoS One. 2018 Jan 2;13(1):e0189688. doi: 10.1371/journal.pone.0189688. eCollection 2018. PLoS One. 2018. PMID: 29293520 Free PMC article.
-
Stimulated and unstimulated saliva samples have significantly different bacterial profiles.PLoS One. 2018 Jun 1;13(6):e0198021. doi: 10.1371/journal.pone.0198021. eCollection 2018. PLoS One. 2018. PMID: 29856779 Free PMC article.
-
Mapping of ribosomal 23S ribosomal RNA modifications in Clostridium sporogenes.RNA Biol. 2018;15(8):1060-1070. doi: 10.1080/15476286.2018.1486662. Epub 2018 Aug 13. RNA Biol. 2018. PMID: 29947286 Free PMC article.
References
-
- Agris PF 1996. The importance of being modified: Roles of modified nucleosides and Mg2+ in RNA structure and function. Prog Nucleic Acid Res Mol Biol 53: 79–129 - PubMed
-
- Agris PF, Vendeix FA, Graham WD 2007. tRNA's wobble decoding of the genome: 40 years of modification. J Mol Biol 366: 1–13 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials