

Structure-based virtual screening of small-molecule antagonists of platelet integrin αIIbβ3 that do not prime the receptor to bind ligand
- PMID: 22893377
- PMCID: PMC3496400
- DOI: 10.1007/s10822-012-9594-6
Structure-based virtual screening of small-molecule antagonists of platelet integrin αIIbβ3 that do not prime the receptor to bind ligand
Abstract
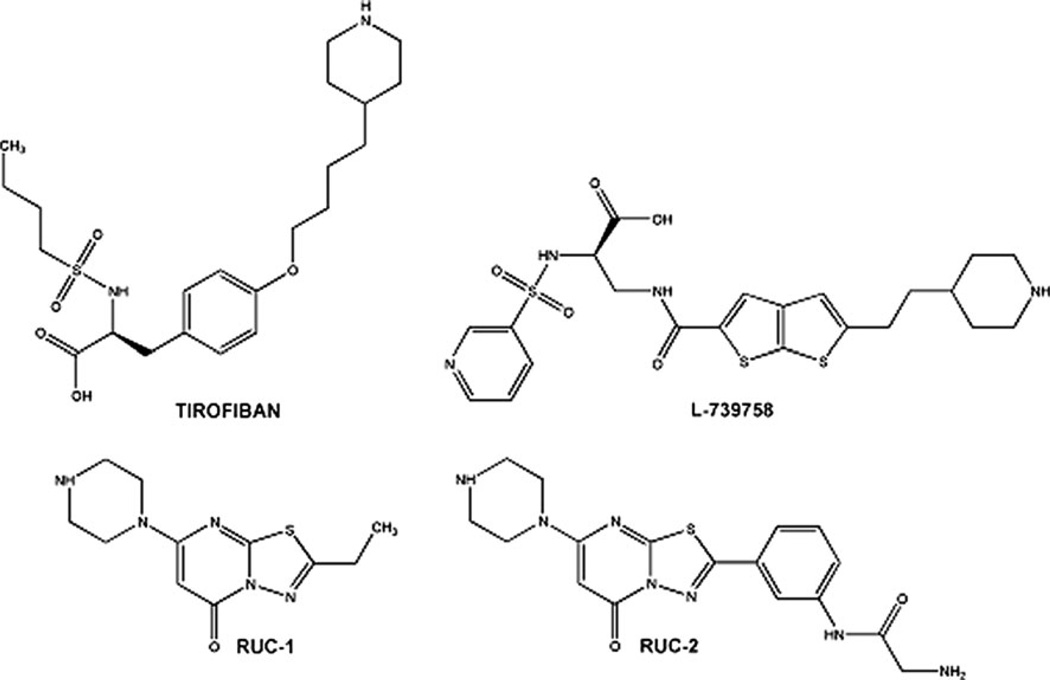
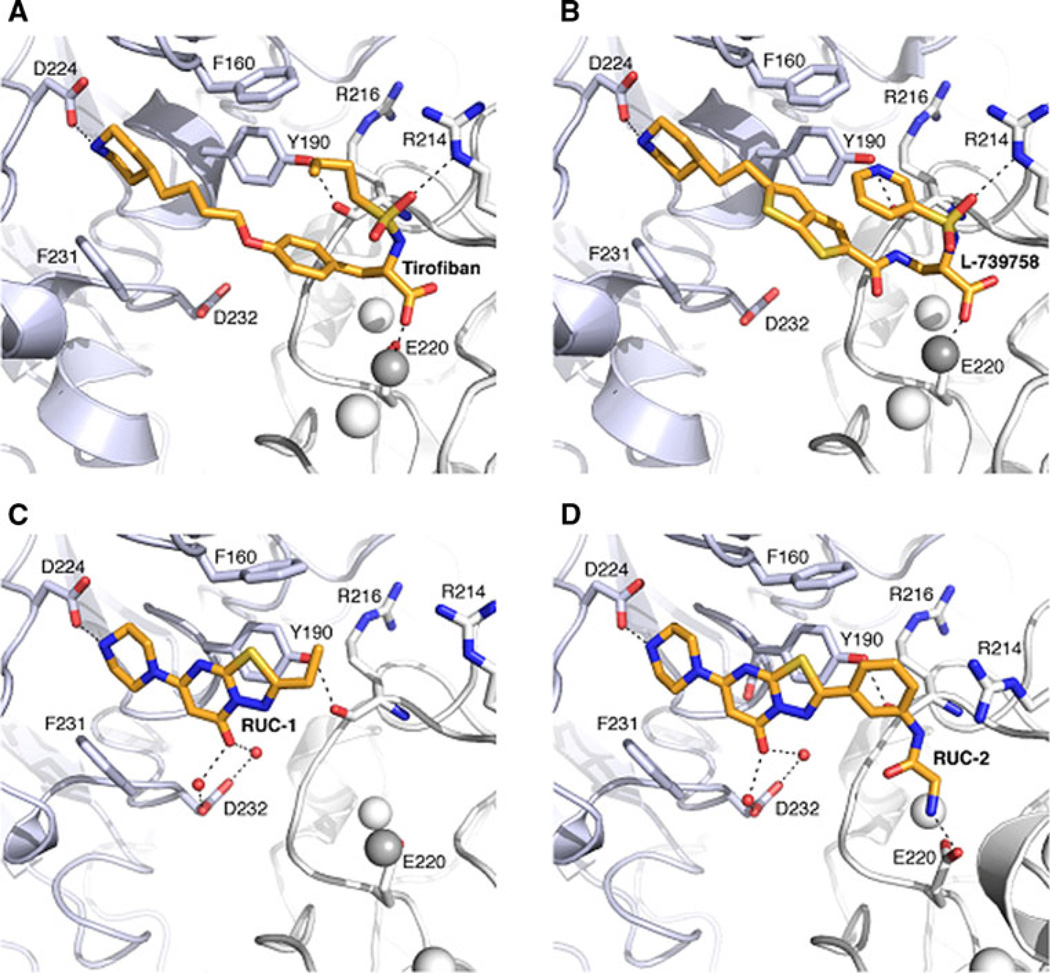
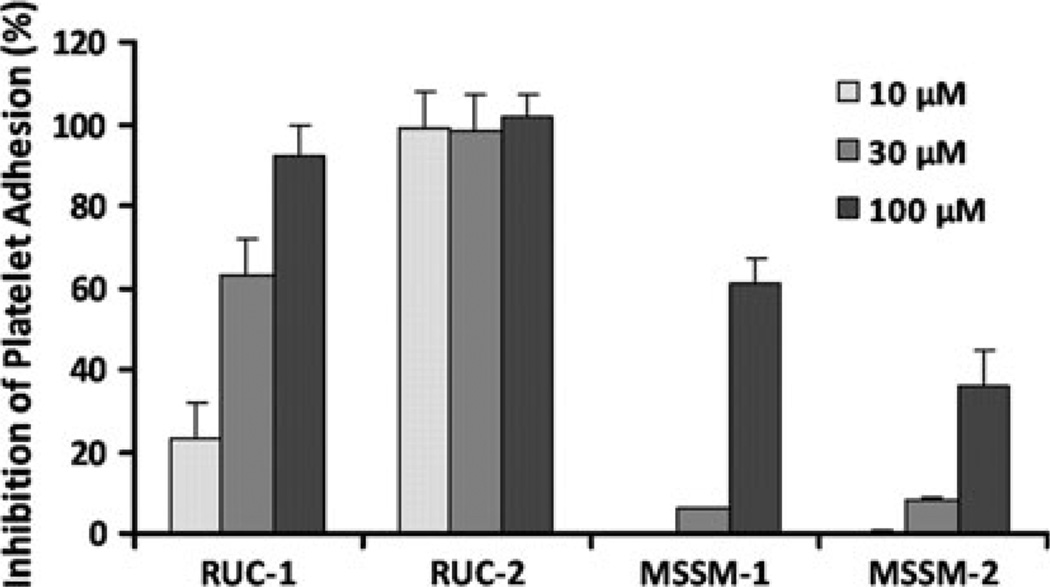
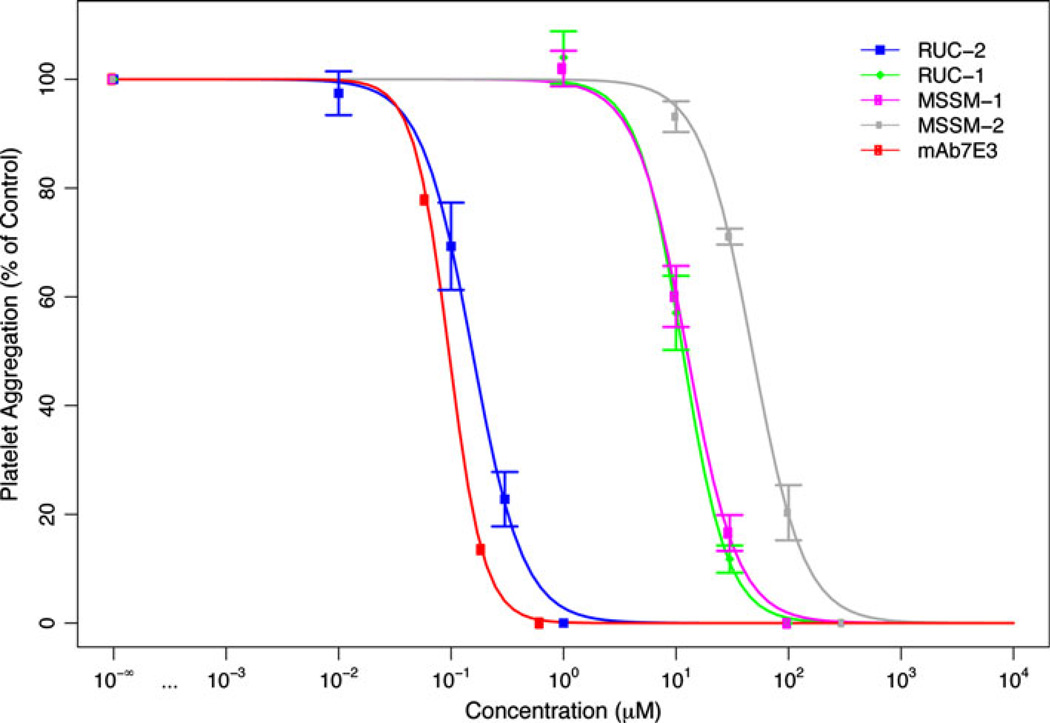
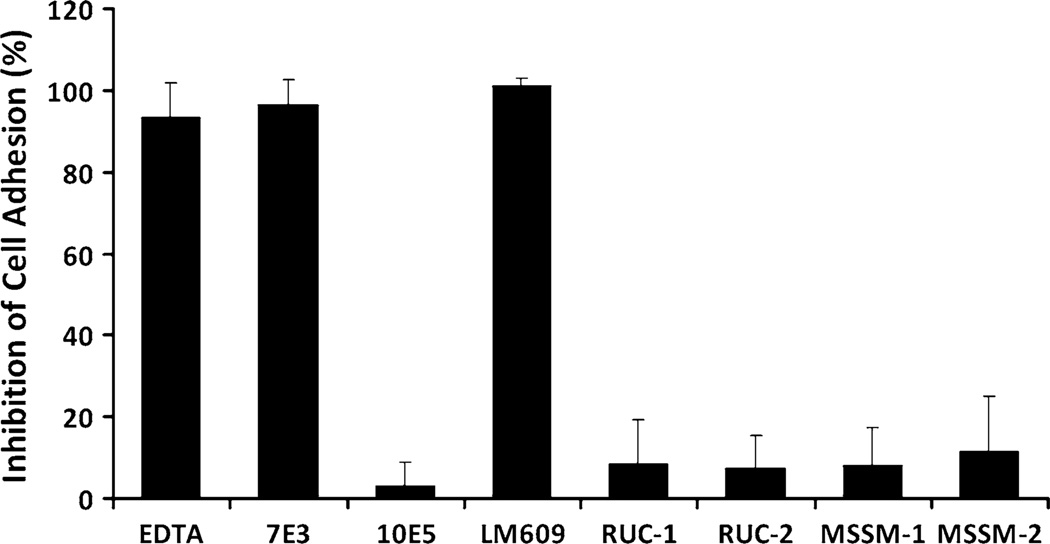
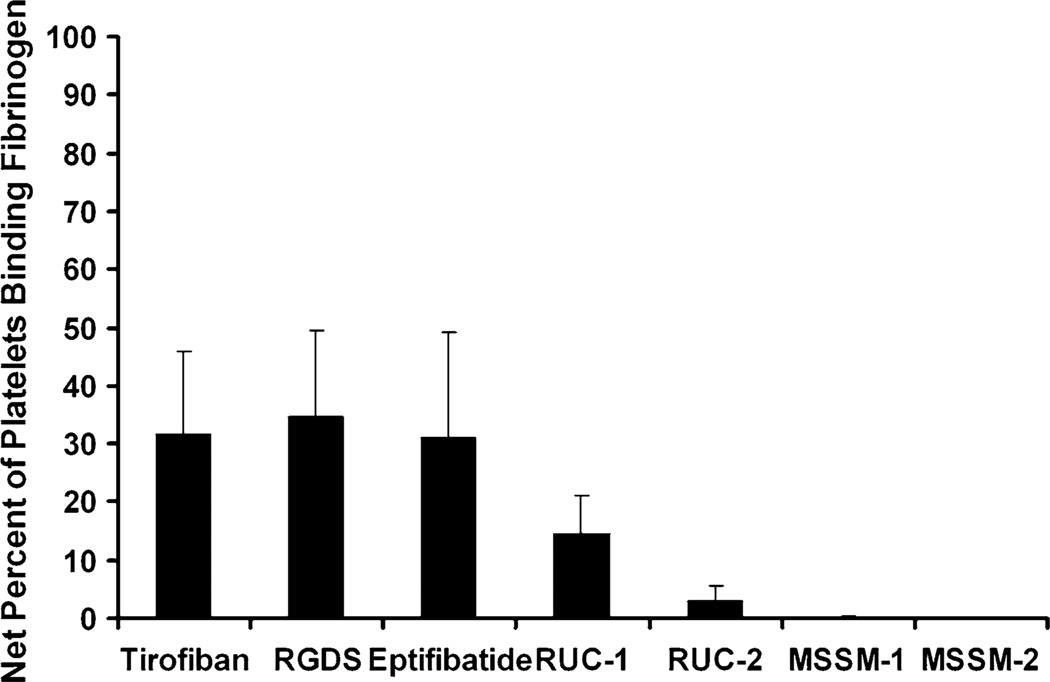
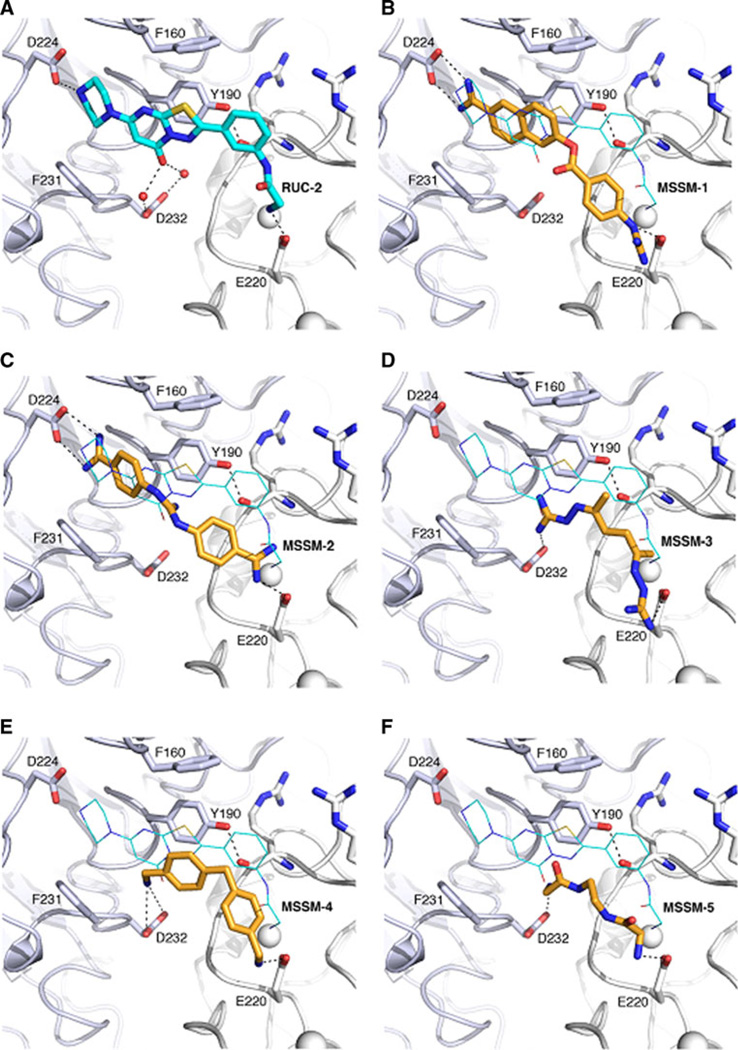
Integrin αIIbβ3 has emerged as an important therapeutic target for thrombotic vascular diseases owing to its pivotal role in mediating platelet aggregation through interaction with adhesive ligands. In the search for effective anti-thrombotic agents that can be administered orally without inducing the high-affinity ligand binding state, we recently discovered via high-throughput screening of 33,264 compounds a novel, αIIbβ3-selective inhibitor (RUC-1) of adenosine-5'-diphosphate (ADP) -induced platelet aggregation that exhibits a different chemical scaffold and mode of binding with respect to classical Arg-Gly-Asp (RGD)-mimicking αIIbβ3 antagonists. Most importantly, RUC-1 and its higher-affinity derivative, RUC-2, do not induce major conformational changes in the protein β3 subunit or prime the receptor to bind ligand. To identify additional αIIbβ3-selective chemotypes that inhibit platelet aggregation through similar mechanisms, we screened in silico over 2.5 million commercially available, 'lead-like' small molecules based on complementarity to the predicted binding mode of RUC-2 into the RUC-1-αIIbβ3 crystal structure. This first reported structure-based virtual screening application to the αIIbβ3 integrin led to the identification of 2 αIIbβ3-selective antagonists out of 4 tested, which compares favorably with the 0.003 % "hit rate" of our previous high-throughput chemical screening study. The newly identified compounds, like RUC-1 and RUC-2, showed specificity for αIIbβ3 compared to αVβ3 and did not prime the receptor to bind ligand. They thus may hold promise as αIIbβ3 antagonist therapeutic scaffolds.
Figures

References
-
- Lopez AD, Mathers CD, Ezzati M, Jamison DT, Murray CJL. Global burden of disease and risk factors, 2011/01/21 edn. Washington: World Bank; 2006. Measuring the global burden of disease and risk factors, 1990–2001. - PubMed
-
- Roger VL, Go AS, Lloyd-Jones DM, Adams RJ, Berry JD, Brown TM, Carnethon MR, Dai S, de Simone G, Ford ES, Fox CS, Fullerton HJ, Gillespie C, Greenlund KJ, Hailpern SM, Heit JA, Ho PM, Howard VJ, Kissela BM, Kittner SJ, Lackland DT, Lichtman JH, Lisabeth LD, Makuc DM, Marcus GM, Marelli A, Matchar DB, McDermott MM, Meigs JB, Moy CS, Mozaffarian D, Mussolino ME, Nichol G, Paynter NP, Rosamond WD, Sorlie PD, Stafford RS, Turan TN, Turner MB, Wong ND, Wylie-Rosett J. Heart disease and stroke statistics—2011 update: a report from the American Heart Association. Circulation. 2011;123(4):e18–e209. - PMC - PubMed
-
- Yang G, Kong L, Zhao W, Wan X, Zhai Y, Chen LC, Koplan JP. Emergence of chronic non-communicable diseases in China. Lancet. 2008;372(9650):1697–1705. - PubMed
-
- Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell. 2002;110(6):673–687. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
