

Poly-N-acetylglucosamine expression by wild-type Yersinia pestis is maximal at mammalian, not flea, temperatures
- PMID: 22893384
- PMCID: PMC3419525
- DOI: 10.1128/mBio.00217-12
Poly-N-acetylglucosamine expression by wild-type Yersinia pestis is maximal at mammalian, not flea, temperatures
Abstract
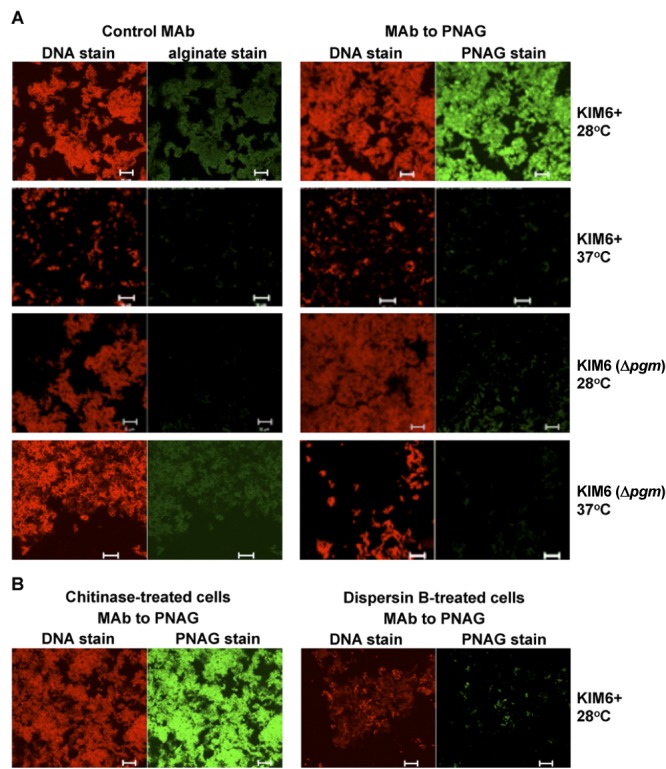
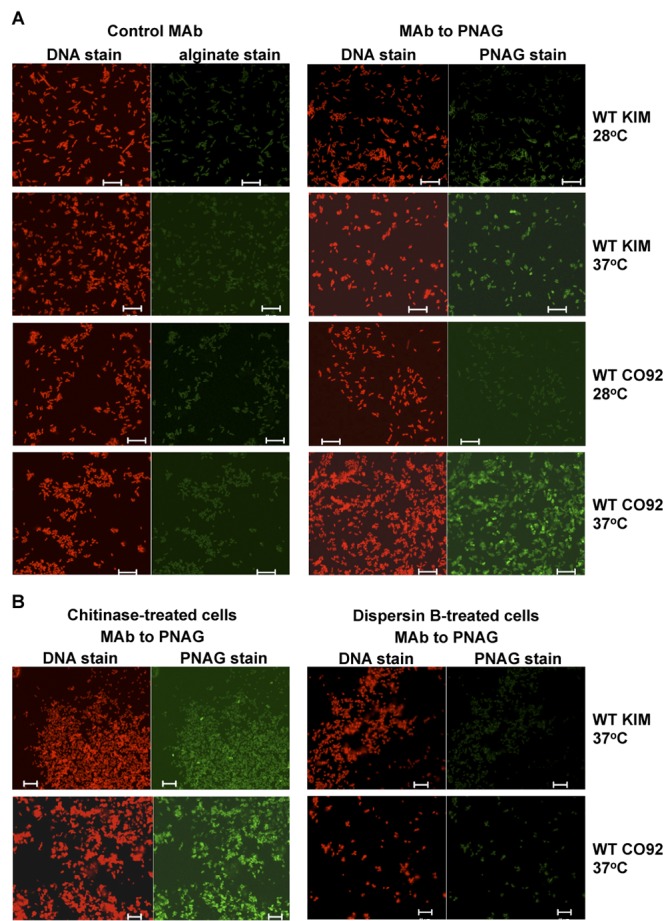
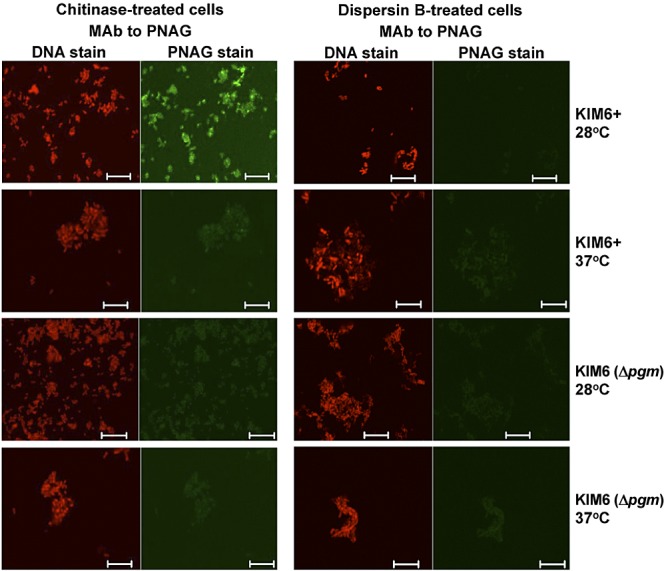
Numerous bacteria, including Yersinia pestis, express the poly-N-acetylglucosamine (PNAG) surface carbohydrate, a major component of biofilms often associated with a specific appearance of colonies on Congo red agar. Biofilm formation and PNAG synthesis by Y. pestis have been reported to be maximal at 21 to 28°C or "flea temperatures," facilitating the regurgitation of Y. pestis into a mammalian host during feeding, but production is diminished at 37°C and thus presumed to be decreased during mammalian infection. Most studies of PNAG expression and biofilm formation by Y. pestis have used a low-virulence derivative of strain KIM, designated KIM6+, that lacks the pCD1 virulence plasmid, and an isogenic mutant without the pigmentation locus, which contains the hemin storage genes that encode PNAG biosynthetic proteins. Using confocal microscopy, fluorescence-activated cell sorter analysis and growth on Congo red agar, we confirmed prior findings regarding PNAG production with the KIM6+ strain. However, we found that fully virulent wild-type (WT) strains KIM and CO92 had maximal PNAG expression at 37°C, with lower PNAG production at 28°C both in broth medium and on Congo red agar plates. Notably, the typical dark colony morphology appearing on Congo red agar was maintained at 28°C, indicating that this phenotype is not associated with PNAG expression in WT Y. pestis. Extracts of WT sylvatic Y. pestis strains from the Russian Federation confirmed the maximal expression of PNAG at 37°C. PNAG production by WT Y. pestis is maximal at mammalian and not insect vector temperatures, suggesting that this factor may have a role during mammalian infection.
Importance: Yersinia pestis transitions from low-temperature residence and replication in insect vectors to higher-temperature replication in mammalian hosts. Prior findings based primarily on an avirulent derivative of WT (wild-type) KIM, named KIM6+, showed that biofilm formation associated with synthesis of poly-N-acetylglucosamine (PNAG) is maximal at 21 to 28°C and decreased at 37°C. Biofilm formation was purported to facilitate the transmission of Y. pestis from fleas to mammals while having little importance in mammalian infection. Here we found that for WT strains KIM and CO92, maximal PNAG production occurs at 37°C, indicating that temperature regulation of PNAG production in WT Y. pestis is not mimicked by strain KIM6+. Additionally, we found that Congo red binding does not always correlate with PNAG production, despite its widespread use as an indicator of biofilm production. Taken together, the findings show that a role for PNAG in WT Y. pestis infection should not be disregarded and warrants further study.
Figures
Similar articles
-
Biovar-related differences apparent in the flea foregut colonization phenotype of distinct Yersinia pestis strains do not impact transmission efficiency.Parasit Vectors. 2020 Jul 1;13(1):335. doi: 10.1186/s13071-020-04207-x. Parasit Vectors. 2020. PMID: 32611387 Free PMC article.
-
Effects of low-temperature flea maintenance on the transmission of Yersinia pestis by Oropsylla montana.Vector Borne Zoonotic Dis. 2013 Jul;13(7):468-78. doi: 10.1089/vbz.2012.1017. Epub 2013 Apr 16. Vector Borne Zoonotic Dis. 2013. PMID: 23590319
-
A Trimeric Autotransporter Enhances Biofilm Cohesiveness in Yersinia pseudotuberculosis but Not in Yersinia pestis.J Bacteriol. 2020 Sep 23;202(20):e00176-20. doi: 10.1128/JB.00176-20. Print 2020 Sep 23. J Bacteriol. 2020. PMID: 32778558 Free PMC article.
-
Yersinia pestis biofilm in the flea vector and its role in the transmission of plague.Curr Top Microbiol Immunol. 2008;322:229-48. doi: 10.1007/978-3-540-75418-3_11. Curr Top Microbiol Immunol. 2008. PMID: 18453279 Free PMC article. Review.
-
Transcriptional regulation of Yersinia pestis biofilm formation.Microb Pathog. 2019 Jun;131:212-217. doi: 10.1016/j.micpath.2019.04.011. Epub 2019 Apr 10. Microb Pathog. 2019. PMID: 30980880 Review.
Cited by
-
Reciprocal regulation of Yersinia pestis biofilm formation and virulence by RovM and RovA.Open Biol. 2016 Mar;6(3):150198. doi: 10.1098/rsob.150198. Open Biol. 2016. PMID: 26984293 Free PMC article.
-
A starvation-induced regulator, RovM, acts as a switch for planktonic/biofilm state transition in Yersinia pseudotuberculosis.Sci Rep. 2017 Apr 4;7(1):639. doi: 10.1038/s41598-017-00534-9. Sci Rep. 2017. PMID: 28377623 Free PMC article.
-
The Cyclic AMP Receptor Protein Regulates Quorum Sensing and Global Gene Expression in Yersinia pestis during Planktonic Growth and Growth in Biofilms.mBio. 2019 Nov 19;10(6):e02613-19. doi: 10.1128/mBio.02613-19. mBio. 2019. PMID: 31744922 Free PMC article.
-
Antibody to a conserved antigenic target is protective against diverse prokaryotic and eukaryotic pathogens.Proc Natl Acad Sci U S A. 2013 Jun 11;110(24):E2209-18. doi: 10.1073/pnas.1303573110. Epub 2013 May 28. Proc Natl Acad Sci U S A. 2013. PMID: 23716675 Free PMC article.
-
Expression and Genetic Activation of Cyclic Di-GMP-Specific Phosphodiesterases in Escherichia coli.J Bacteriol. 2015 Nov 9;198(3):448-62. doi: 10.1128/JB.00604-15. Print 2016 Feb 1. J Bacteriol. 2015. PMID: 26553851 Free PMC article.
References
-
- Jarrett CO, et al. 2004. Transmission of Yersinia pestis from an infectious biofilm in the flea vector. J. Infect. Dis. 190:783–792 - PubMed
-
- Hinnebusch BJ, Perry RD, Schwan TG. 1996. Role of the Yersinia pestis hemin storage (hms) locus in the transmission of plague by fleas. Science 273:367–370 - PubMed
-
- Jones HA, Lillard JW, Jr, Perry RD. 1999. HmsT, a protein essential for essential of the haemin storage (Hms+) phenotype of Yersinia pestis. Microbiology 145:2117–2128 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
