

Prion pathogenesis and secondary lymphoid organs (SLO): tracking the SLO spread of prions to the brain
- PMID: 22895090
- PMCID: PMC3609058
- DOI: 10.4161/pri.20676
Prion pathogenesis and secondary lymphoid organs (SLO): tracking the SLO spread of prions to the brain
Abstract
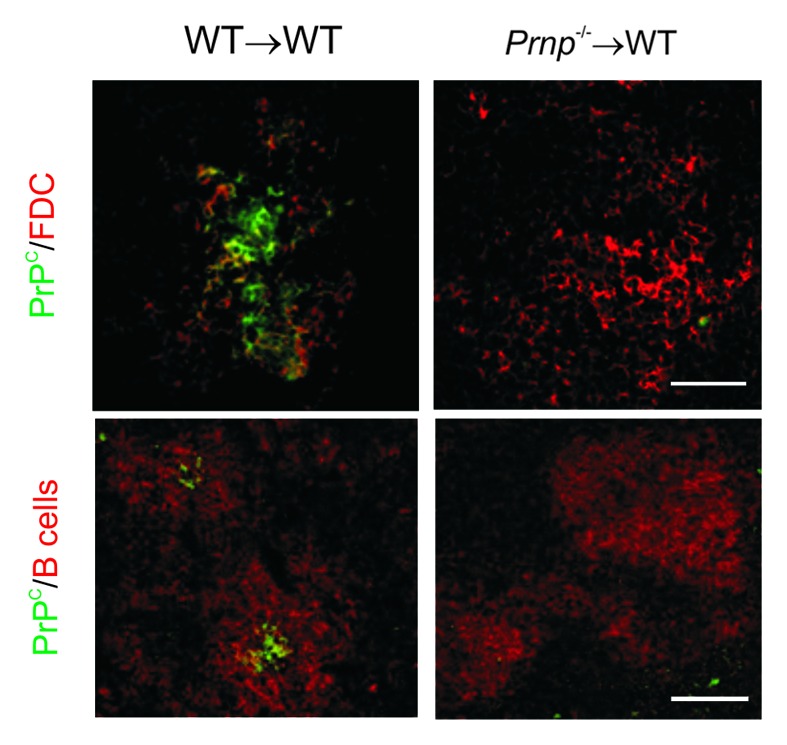
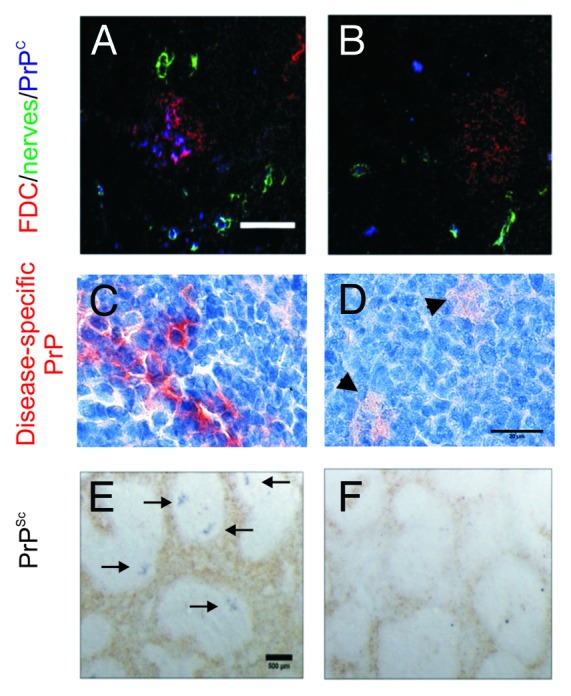
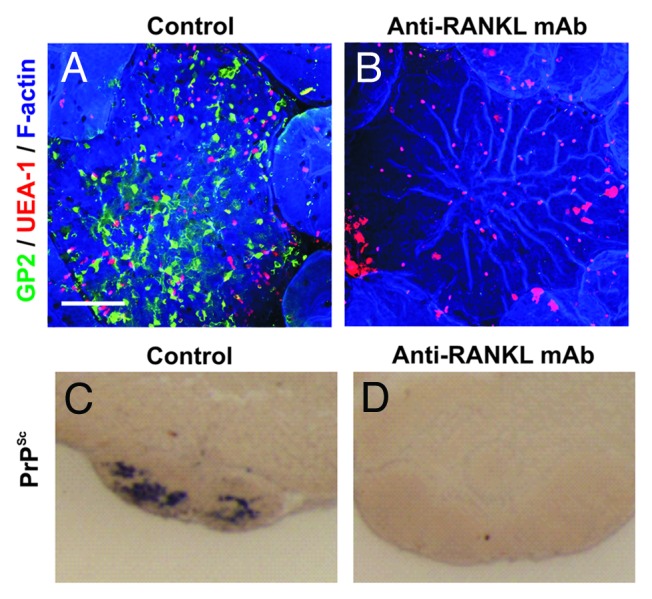
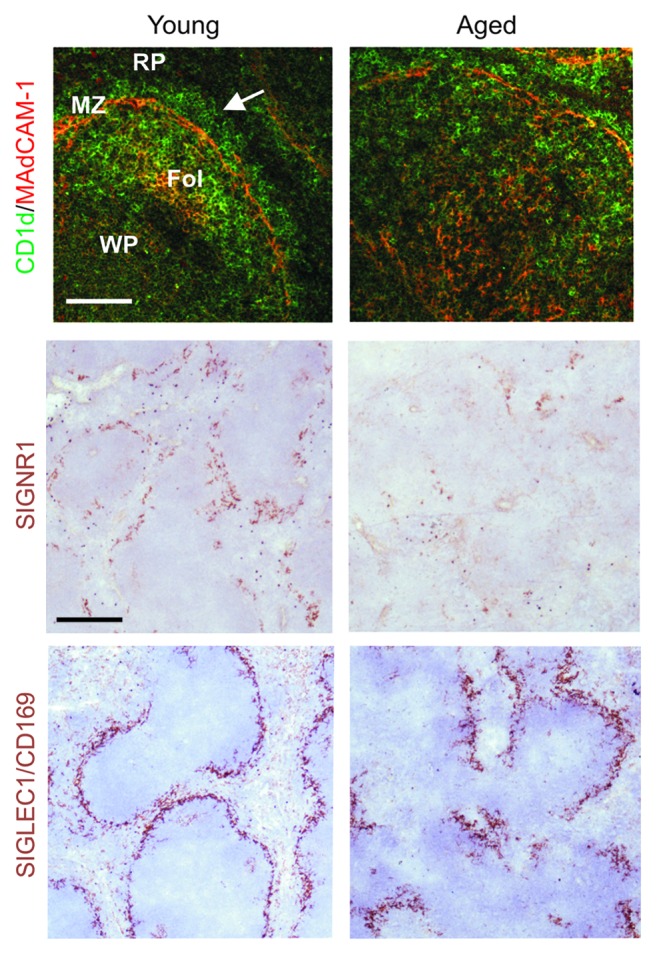
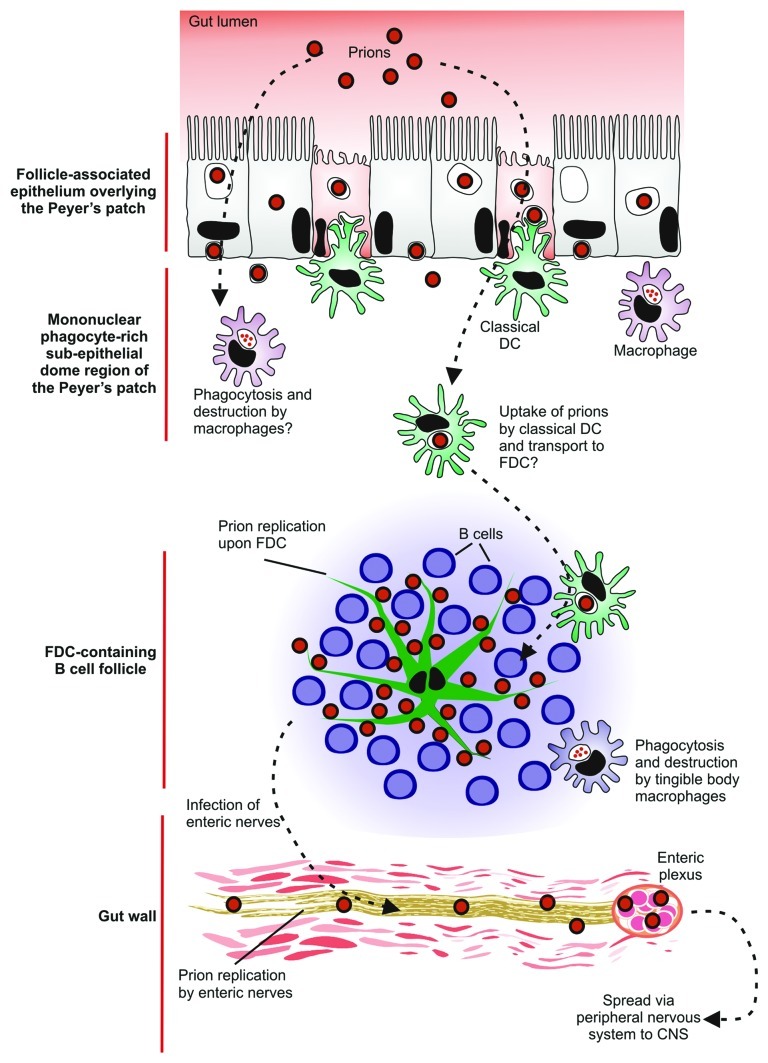
Prion diseases are subacute neurodegenerative diseases that affect humans and a range of domestic and free-ranging animal species. These diseases are characterized by the accumulation of PrP (Sc), an abnormally folded isoform of the cellular prion protein (PrP (C)), in affected tissues. The pathology during prion disease appears to occur almost exclusively within the central nervous system. The extensive neurodegeneration which occurs ultimately leads to the death of the host. An intriguing feature of the prion diseases, when compared with other protein-misfolding diseases, is their transmissibility. Following peripheral exposure, some prion diseases accumulate to high levels within lymphoid tissues. The replication of prions within lymphoid tissue has been shown to be important for the efficient spread of disease to the brain. This article describes recent progress in our understanding of the cellular mechanisms that influence the propagation of prions from peripheral sites of exposure (such as the lumen of the intestine) to the brain. A thorough understanding of these events will lead to the identification of important targets for therapeutic intervention, or alternatively, reveal additional processes that influence disease susceptibility to peripherally-acquired prion diseases.
Figures
Similar articles
-
How do PrPSc Prions Spread between Host Species, and within Hosts?Pathogens. 2017 Nov 24;6(4):60. doi: 10.3390/pathogens6040060. Pathogens. 2017. PMID: 29186791 Free PMC article. Review.
-
Oral Prion Neuroinvasion Occurs Independently of PrPC Expression in the Gut Epithelium.J Virol. 2018 Sep 12;92(19):e01010-18. doi: 10.1128/JVI.01010-18. Print 2018 Oct 1. J Virol. 2018. PMID: 30021891 Free PMC article.
-
The role of the immune system in prion infection.Handb Clin Neurol. 2018;153:85-107. doi: 10.1016/B978-0-444-63945-5.00005-2. Handb Clin Neurol. 2018. PMID: 29887157 Review.
-
Immunology of Prion Protein and Prions.Prog Mol Biol Transl Sci. 2017;150:203-240. doi: 10.1016/bs.pmbts.2017.06.004. Epub 2017 Jul 31. Prog Mol Biol Transl Sci. 2017. PMID: 28838662 Review.
-
Follicular dendritic cell-specific prion protein (PrP) expression alone is sufficient to sustain prion infection in the spleen.PLoS Pathog. 2011 Dec;7(12):e1002402. doi: 10.1371/journal.ppat.1002402. Epub 2011 Dec 1. PLoS Pathog. 2011. PMID: 22144895 Free PMC article.
Cited by
-
Prion pathogenesis is unaltered following down-regulation of SIGN-R1.Virology. 2016 Oct;497:337-345. doi: 10.1016/j.virol.2016.08.005. Epub 2016 Aug 11. Virology. 2016. PMID: 27522473 Free PMC article.
-
Appendectomy, Tonsillectomy and Parkinson's Disease Risk: A Swedish Register-Based Study.Front Neurol. 2020 Jun 5;11:510. doi: 10.3389/fneur.2020.00510. eCollection 2020. Front Neurol. 2020. PMID: 32595591 Free PMC article.
-
Environmental and host factors that contribute to prion strain evolution.Acta Neuropathol. 2021 Jul;142(1):5-16. doi: 10.1007/s00401-021-02310-6. Epub 2021 Apr 25. Acta Neuropathol. 2021. PMID: 33899132 Free PMC article. Review.
-
Cases of Creutzfeldt-Jakob disease in young individuals: open questions regarding aetiology.Front Cell Neurosci. 2025 Apr 14;19:1571662. doi: 10.3389/fncel.2025.1571662. eCollection 2025. Front Cell Neurosci. 2025. PMID: 40297707 Free PMC article. No abstract available.
-
Limited understanding of the functional diversity of N-linked glycans as a major gap of prion biology.Prion. 2017 Mar 4;11(2):82-88. doi: 10.1080/19336896.2017.1301338. Epub 2017 Mar 21. Prion. 2017. PMID: 28324664 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- BB/D00831X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/D/20251967/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/A/00001660/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/A/00001659/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MRC_/Medical Research Council/United Kingdom
- BB/D00831X/3/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/G003947/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/D/20251968/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/D/05241339/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/D00831X/2/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F019726/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/I/00000989/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Research Materials