

Bidirectional plasticity of intrinsic excitability controls sensory inputs efficiency in layer 5 barrel cortex neurons in vivo
- PMID: 22895720
- PMCID: PMC6621180
- DOI: 10.1523/JNEUROSCI.0415-12.2012
Bidirectional plasticity of intrinsic excitability controls sensory inputs efficiency in layer 5 barrel cortex neurons in vivo
Abstract
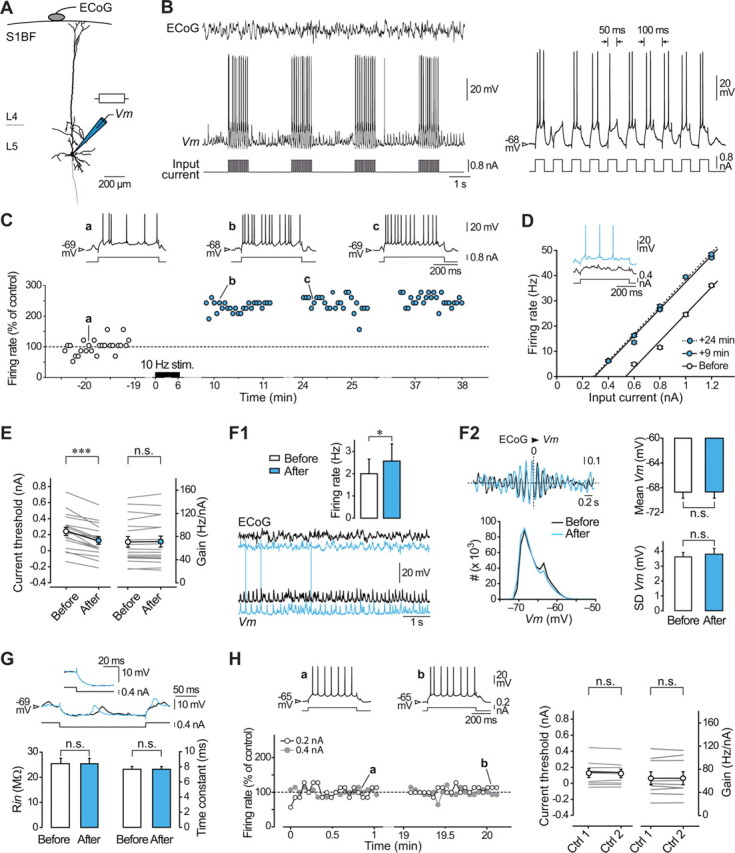
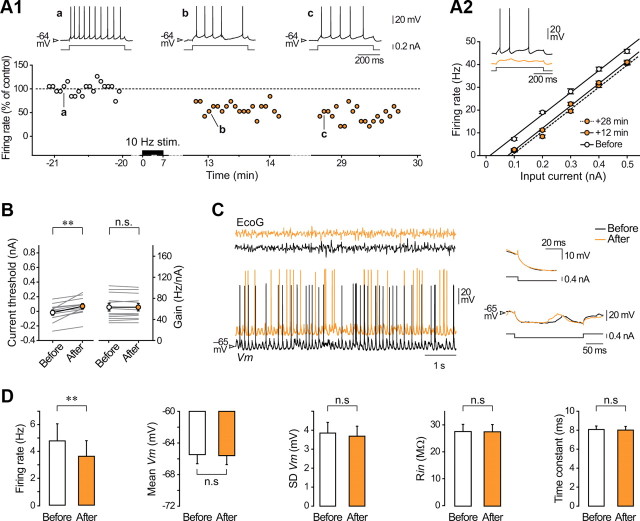
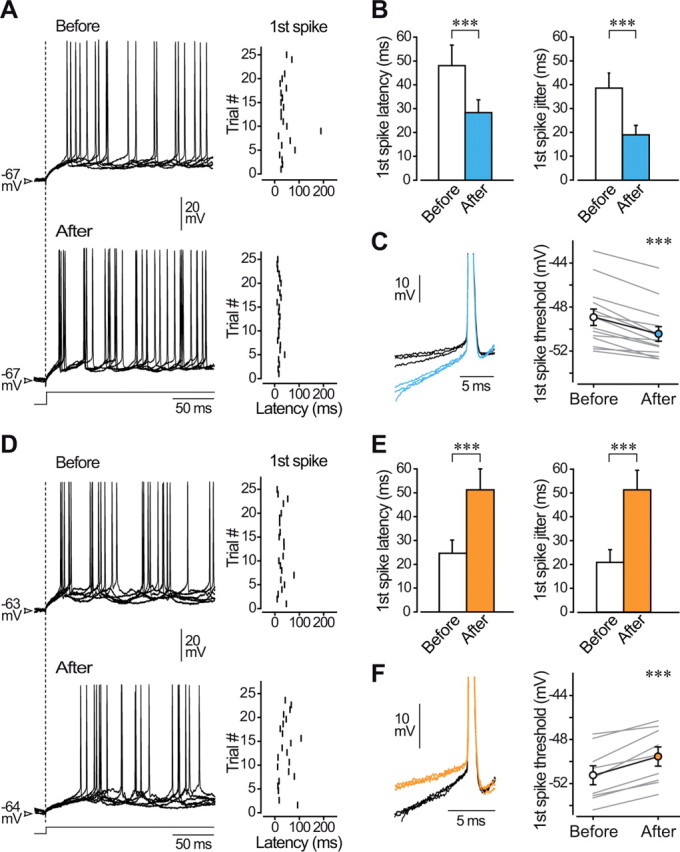
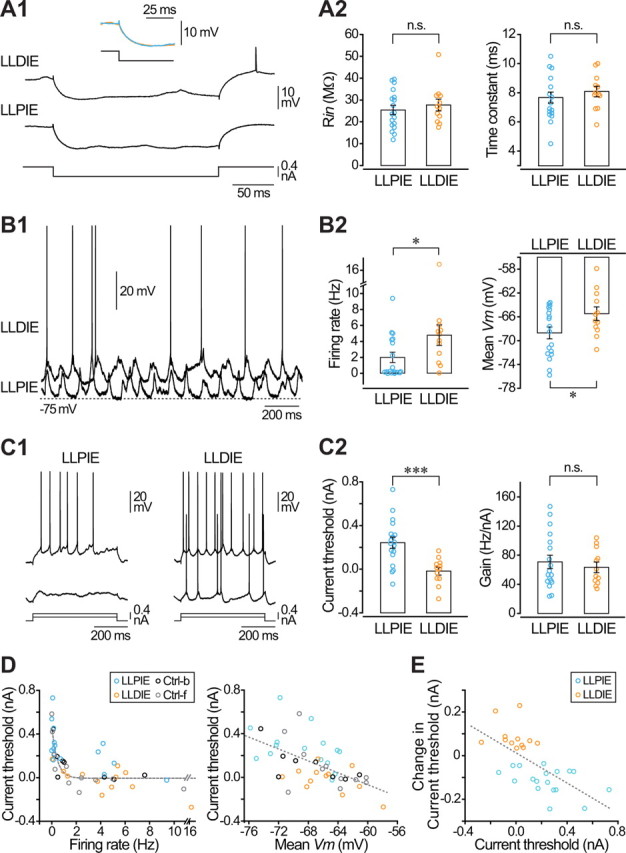
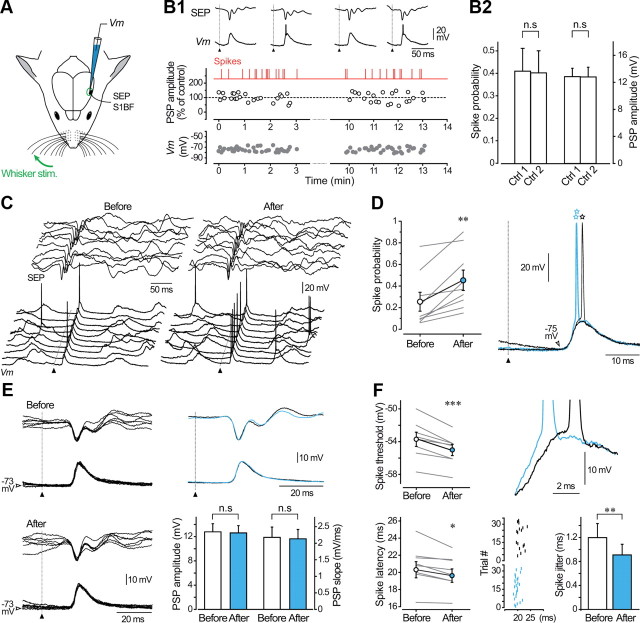
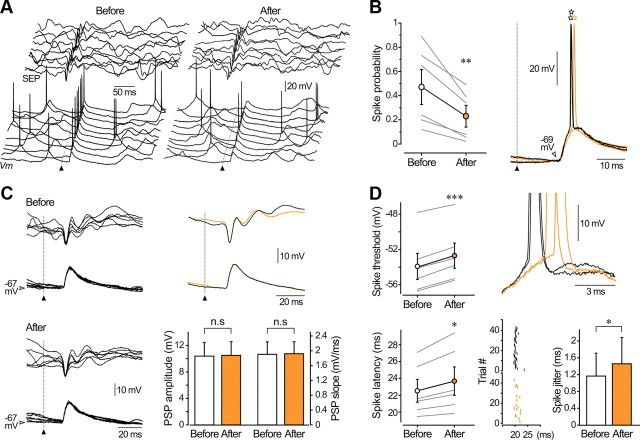
Responsiveness of cortical neurons to sensory inputs can be altered by experience and learning. While synaptic plasticity is generally proposed as the underlying cellular mechanism, possible contributions of activity-dependent changes in intrinsic excitability remain poorly investigated. Here, we show that periods of rhythmic firing in rat barrel cortex layer 5 pyramidal neurons can trigger a long-lasting increase or decrease in their membrane excitability in vivo. Potentiation of cortical excitability consisted of an increased firing in response to intracellular stimulation and a reduction in threshold current for spike initiation. Conversely, depression of cortical excitability was evidenced by an augmented firing threshold leading to a reduced current-evoked spiking. The direction of plasticity depended on the baseline level of spontaneous firing rate and cell excitability. We also found that changes in intrinsic excitability were accompanied by corresponding modifications in the effectiveness of sensory inputs. Potentiation and depression of cortical neuron excitability resulted, respectively, in an increased or decreased firing probability on whisker-evoked synaptic responses, without modifications in the synaptic strength itself. These data suggest that bidirectional intrinsic plasticity could play an important role in experience-dependent refinement of sensory cortical networks.
Figures
Similar articles
-
Whisker row deprivation affects the flow of sensory information through rat barrel cortex.J Neurophysiol. 2017 Jan 1;117(1):4-17. doi: 10.1152/jn.00289.2016. Epub 2016 Oct 5. J Neurophysiol. 2017. PMID: 27707809 Free PMC article.
-
Chronic suppression of activity in barrel field cortex downregulates sensory responses in contralateral barrel field cortex.J Neurophysiol. 2005 Nov;94(5):3342-56. doi: 10.1152/jn.00357.2005. Epub 2005 Jul 13. J Neurophysiol. 2005. PMID: 16014795
-
Rapid Disinhibition by Adjustment of PV Intrinsic Excitability during Whisker Map Plasticity in Mouse S1.J Neurosci. 2018 May 16;38(20):4749-4761. doi: 10.1523/JNEUROSCI.3628-17.2018. Epub 2018 Apr 20. J Neurosci. 2018. PMID: 29678876 Free PMC article.
-
From single cells and single columns to cortical networks: dendritic excitability, coincidence detection and synaptic transmission in brain slices and brains.Exp Physiol. 2017 May 1;102(5):489-521. doi: 10.1113/EP085776. Epub 2017 Apr 21. Exp Physiol. 2017. PMID: 28139019 Free PMC article. Review.
-
A comparison of experience-dependent plasticity in the visual and somatosensory systems.Neuron. 2005 Nov 3;48(3):465-77. doi: 10.1016/j.neuron.2005.10.013. Neuron. 2005. PMID: 16269363 Review.
Cited by
-
Sensory coding is impaired in rat absence epilepsy.J Physiol. 2019 Feb;597(3):951-966. doi: 10.1113/JP277297. Epub 2019 Jan 4. J Physiol. 2019. PMID: 30548850 Free PMC article.
-
Toward a Neurocentric View of Learning.Neuron. 2017 Jul 5;95(1):19-32. doi: 10.1016/j.neuron.2017.05.021. Neuron. 2017. PMID: 28683265 Free PMC article. Review.
-
Intrinsic plasticity and birdsong learning.Neurobiol Learn Mem. 2021 Apr;180:107407. doi: 10.1016/j.nlm.2021.107407. Epub 2021 Feb 22. Neurobiol Learn Mem. 2021. PMID: 33631346 Free PMC article. Review.
-
Microglia-triggered hypoexcitability plasticity of pyramidal neurons in the rat medial prefrontal cortex.Curr Res Neurobiol. 2022 Feb 5;3:100028. doi: 10.1016/j.crneur.2022.100028. eCollection 2022. Curr Res Neurobiol. 2022. PMID: 36518339 Free PMC article.
-
The contribution of low contrast-preferring neurons to information representation in the primary visual cortex after learning.Sci Adv. 2021 Nov 26;7(48):eabj9976. doi: 10.1126/sciadv.abj9976. Epub 2021 Nov 26. Sci Adv. 2021. PMID: 34826242 Free PMC article.
References
-
- Aizenman CD, Akerman CJ, Jensen KR, Cline HT. Visually driven regulation of intrinsic neuronal excitability improves stimulus detection in vivo. Neuron. 2003;39:831–842. - PubMed
-
- Bekisz M, Garkun Y, Wabno J, Hess G, Wrobel A, Kossut M. Increased excitability of cortical neurons induced by associative learning: an ex vivo study. Eur J Neurosci. 2010;32:1715–1725. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources