

Response properties of local field potentials and neighboring single neurons in awake primary visual cortex
- PMID: 22895722
- PMCID: PMC3436073
- DOI: 10.1523/JNEUROSCI.0429-12.2012
Response properties of local field potentials and neighboring single neurons in awake primary visual cortex
Abstract
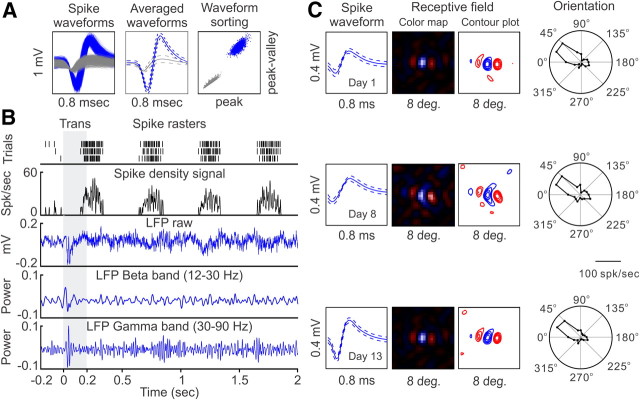
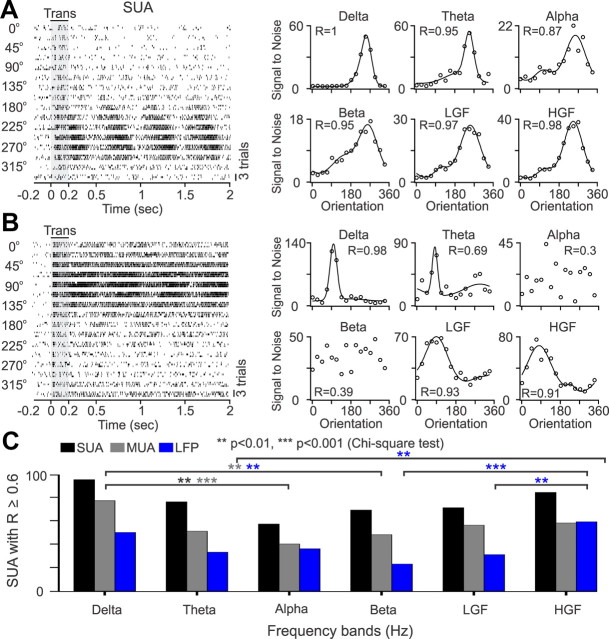
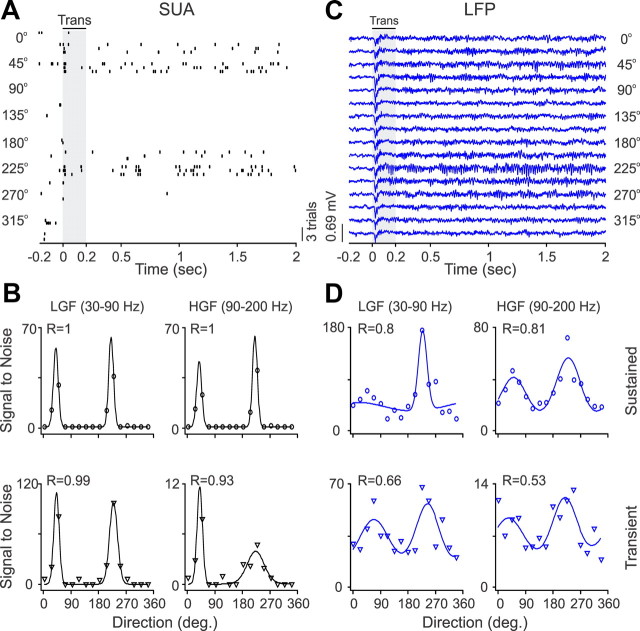
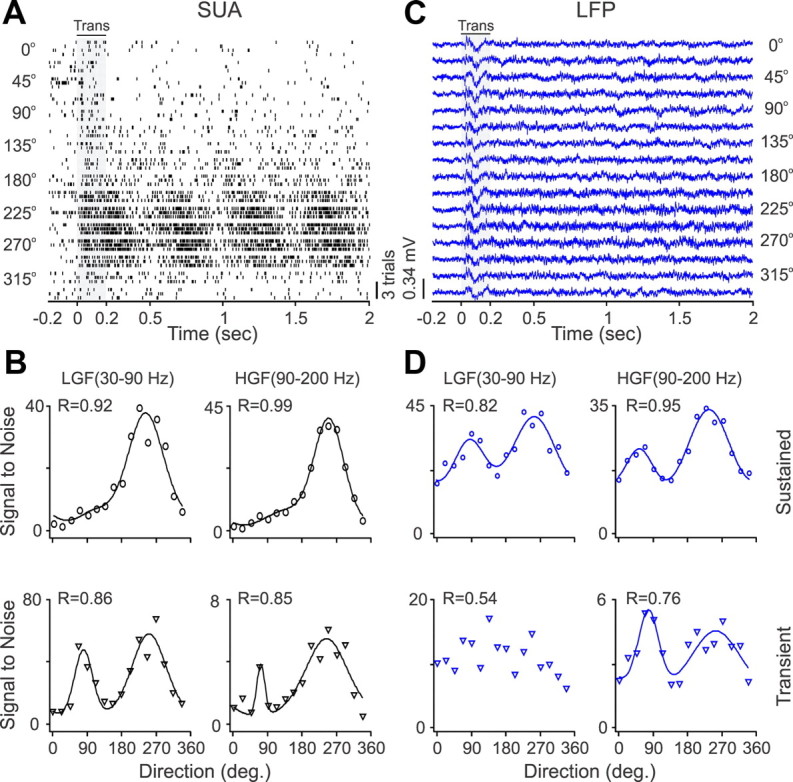
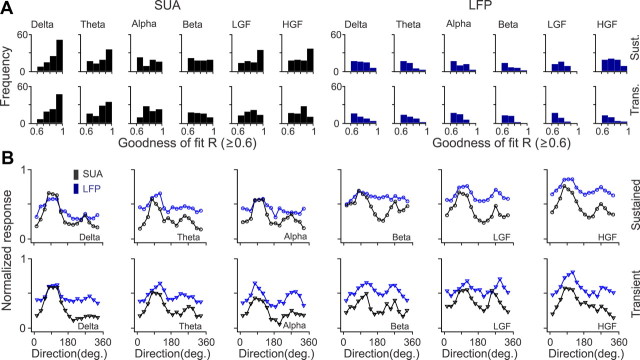
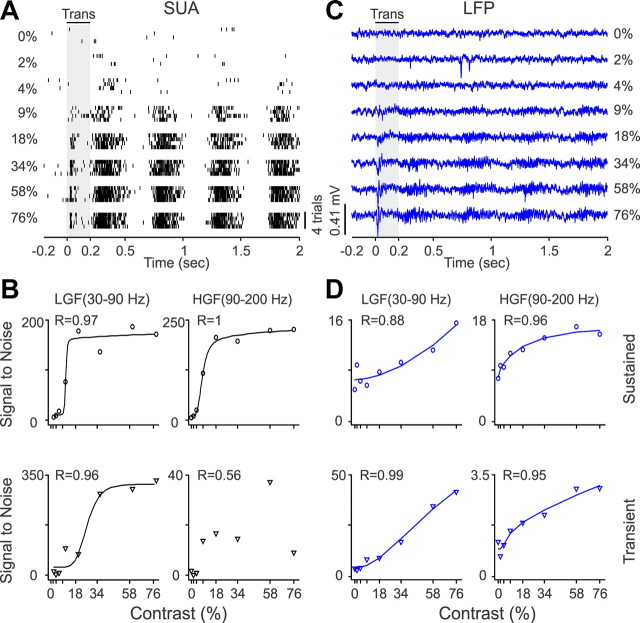
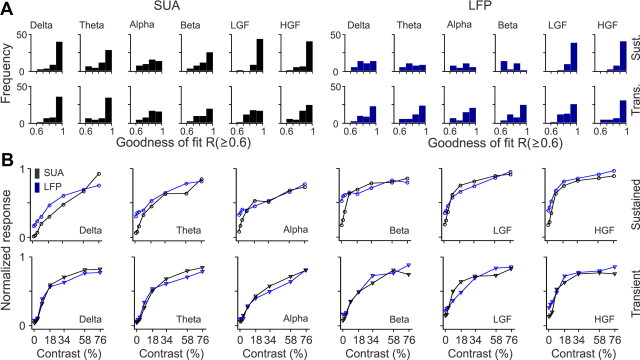
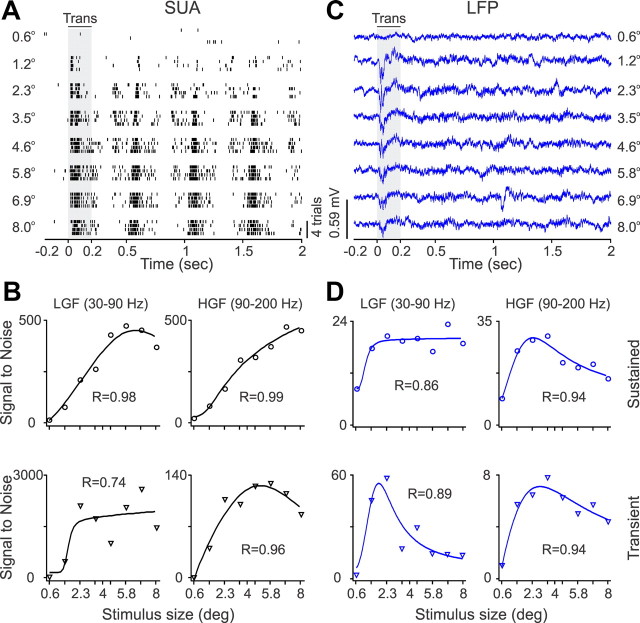
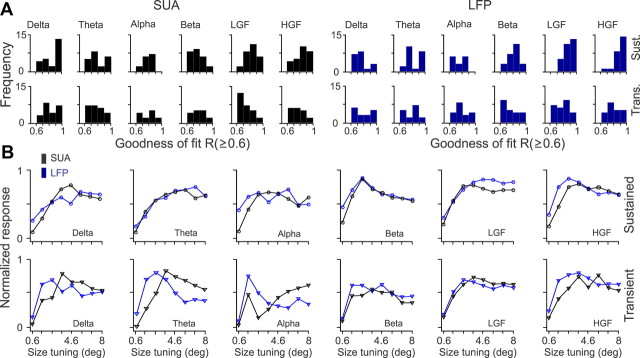
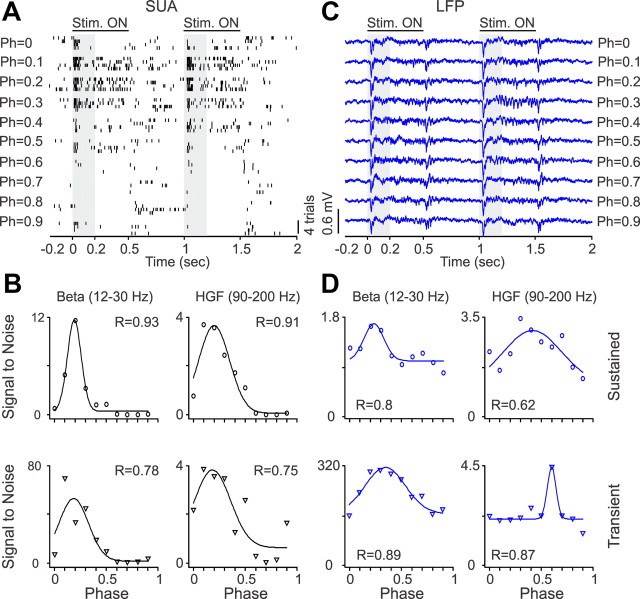
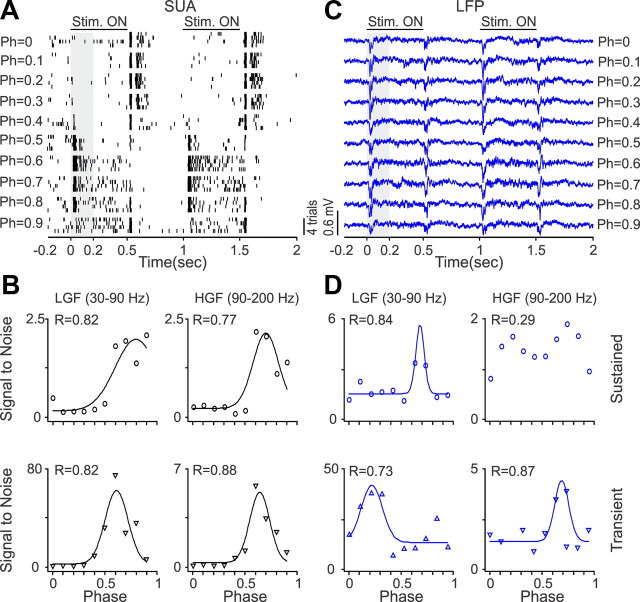
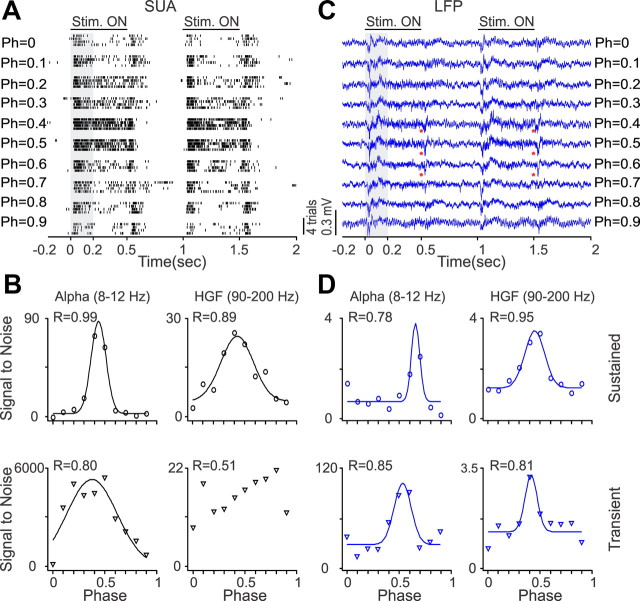
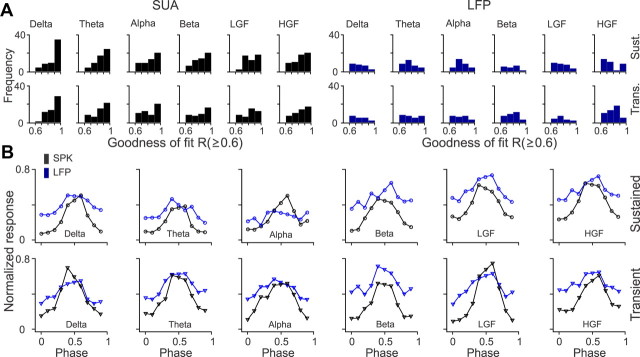
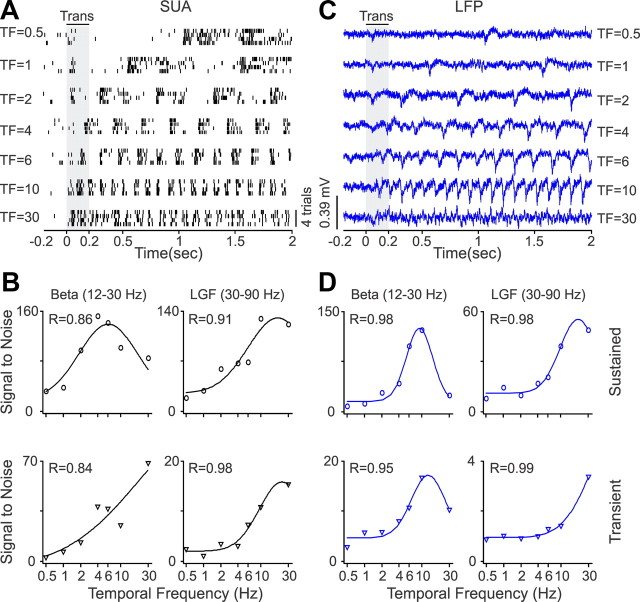
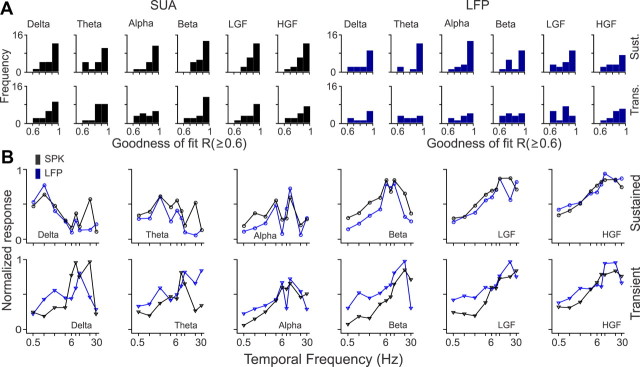
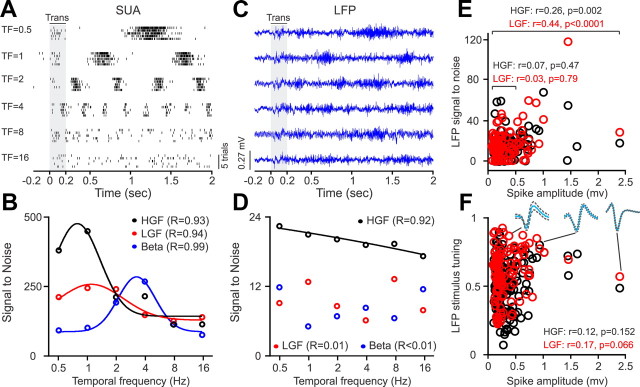
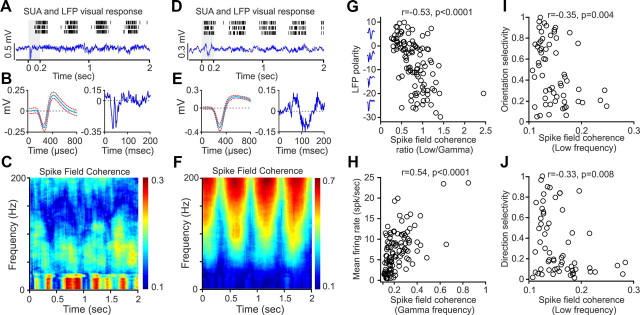
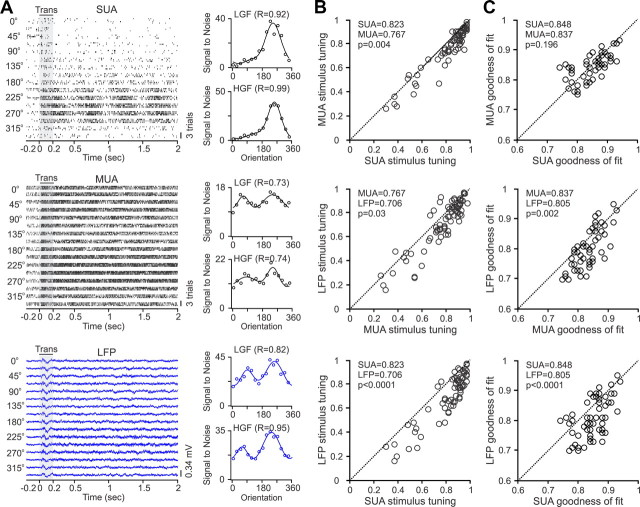
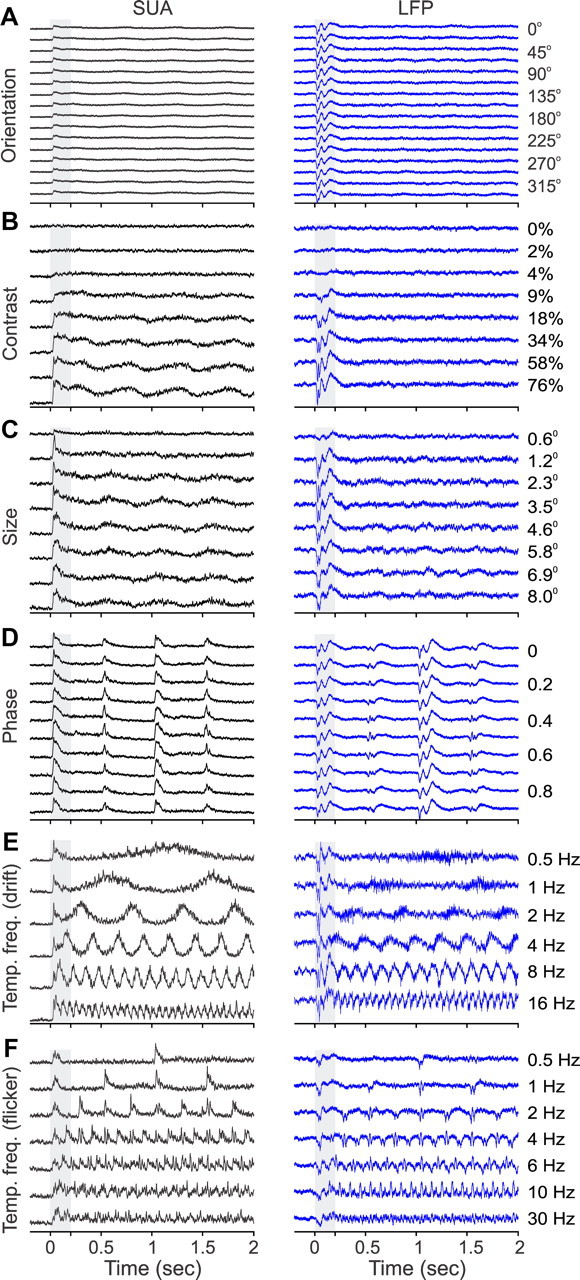
Recordings from local field potentials (LFPs) are becoming increasingly common in research and clinical applications, but we still have a poor understanding of how LFP stimulus selectivity originates from the combined activity of single neurons. Here, we systematically compared the stimulus selectivity of LFP and neighboring single-unit activity (SUA) recorded in area primary visual cortex (V1) of awake primates. We demonstrate that LFP and SUA have similar stimulus preferences for orientation, direction of motion, contrast, size, temporal frequency, and even spatial phase. However, the average SUA had 50 times better signal-to-noise, 20% higher contrast sensitivity, 45% higher direction selectivity, and 15% more tuning depth than the average LFP. Low LFP frequencies (<30 Hz) were most strongly correlated with the spiking frequencies of neurons with nonlinear spatial summation and poor orientation/direction selectivity that were located near cortical current sinks (negative LFPs). In contrast, LFP gamma frequencies (>30 Hz) were correlated with a more diverse group of neurons located near cortical sources (positive LFPs). In summary, our results indicate that low- and high-frequency LFP pool signals from V1 neurons with similar stimulus preferences but different response properties and cortical depths.
Figures
Similar articles
-
The local and non-local components of the local field potential in awake primate visual cortex.J Comput Neurosci. 2010 Dec;29(3):615-23. doi: 10.1007/s10827-010-0223-x. Epub 2010 Feb 24. J Comput Neurosci. 2010. PMID: 20180148
-
Chromatic and Achromatic Spatial Resolution of Local Field Potentials in Awake Cortex.Cereb Cortex. 2015 Oct;25(10):3877-93. doi: 10.1093/cercor/bhu270. Epub 2014 Nov 21. Cereb Cortex. 2015. PMID: 25416722 Free PMC article.
-
Transitions between Multiband Oscillatory Patterns Characterize Memory-Guided Perceptual Decisions in Prefrontal Circuits.J Neurosci. 2016 Jan 13;36(2):489-505. doi: 10.1523/JNEUROSCI.3678-15.2016. J Neurosci. 2016. PMID: 26758840 Free PMC article.
-
Modelling and analysis of local field potentials for studying the function of cortical circuits.Nat Rev Neurosci. 2013 Nov;14(11):770-85. doi: 10.1038/nrn3599. Nat Rev Neurosci. 2013. PMID: 24135696 Review.
-
Specificity and randomness in the visual cortex.Curr Opin Neurobiol. 2007 Aug;17(4):401-7. doi: 10.1016/j.conb.2007.07.007. Epub 2007 Aug 27. Curr Opin Neurobiol. 2007. PMID: 17720489 Free PMC article. Review.
Cited by
-
Salience of unique hues and implications for color theory.J Vis. 2015 Feb 6;15(2):10. doi: 10.1167/15.2.10. J Vis. 2015. PMID: 25761328 Free PMC article.
-
LFP polarity changes across cortical and eccentricity in primary visual cortex.Front Neurosci. 2023 Feb 27;17:1138602. doi: 10.3389/fnins.2023.1138602. eCollection 2023. Front Neurosci. 2023. PMID: 36922925 Free PMC article.
-
The Primary Visual Cortex Is Differentially Modulated by Stimulus-Driven and Top-Down Attention.PLoS One. 2016 Jan 5;11(1):e0145379. doi: 10.1371/journal.pone.0145379. eCollection 2016. PLoS One. 2016. PMID: 26730705 Free PMC article.
-
Distinct frequency bands in the local field potential are differently tuned to stimulus drift rate.J Neurophysiol. 2018 Aug 1;120(2):681-692. doi: 10.1152/jn.00807.2017. Epub 2018 Apr 25. J Neurophysiol. 2018. PMID: 29694281 Free PMC article.
-
Online prediction of optimal deep brain stimulation contacts from local field potentials in Parkinson's disease.NPJ Parkinsons Dis. 2025 Aug 8;11(1):234. doi: 10.1038/s41531-025-01092-y. NPJ Parkinsons Dis. 2025. PMID: 40781085 Free PMC article.
References
-
- Albright TD, Desimone R, Gross CG. Columnar organization of directionally selective cells in visual area MT of the macaque. J Neurophysiol. 1984;51:16–31. - PubMed
-
- Albus K. A quantitative study of the projection area of the central and the paracentral visual field in area 17 of the cat. I. The precision of the topography. Exp Brain Res. 1975;24:159–179. - PubMed
-
- Andersen RA, Musallam S, Pesaran B. Selecting the signals for a brain-machine interface. Curr Opin Neurobiol. 2004;14:720–726. - PubMed
-
- Aronov D, Reich DS, Mechler F, Victor JD. Neural coding of spatial phase in V1 of the macaque monkey. J Neurophysiol. 2003;89:3304–3327. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources