

Rate limit of protein elastic response is tether dependent
- PMID: 22895787
- PMCID: PMC3437906
- DOI: 10.1073/pnas.1212167109
Rate limit of protein elastic response is tether dependent
Abstract
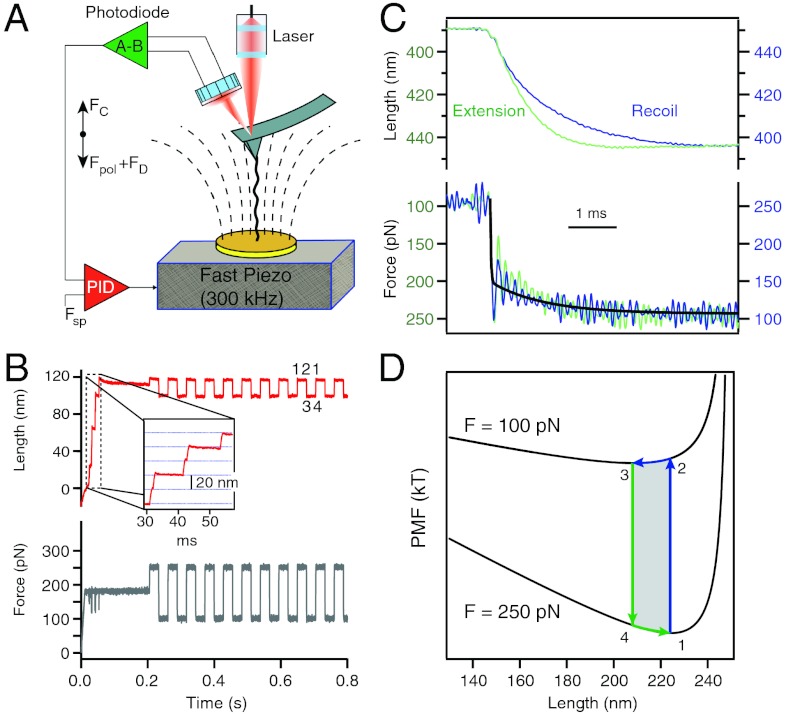
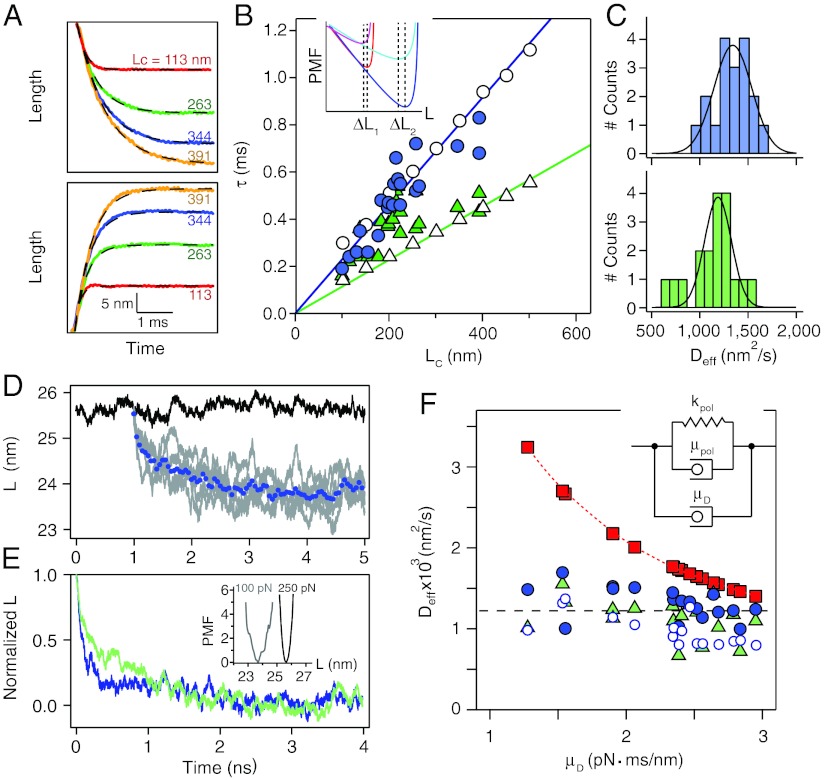
The elastic restoring force of tissues must be able to operate over the very wide range of loading rates experienced by living organisms. It is surprising that even the fastest events involving animal muscle tissues do not surpass a few hundred hertz. We propose that this limit is set in part by the elastic dynamics of tethered proteins extending and relaxing under a changing load. Here we study the elastic dynamics of tethered proteins using a fast force spectrometer with sub-millisecond time resolution, combined with Brownian and Molecular Dynamics simulations. We show that the act of tethering a polypeptide to an object, an inseparable part of protein elasticity in vivo and in experimental setups, greatly reduces the attempt frequency with which the protein samples its free energy. Indeed, our data shows that a tethered polypeptide can traverse its free-energy landscape with a surprisingly low effective diffusion coefficient D(eff) ~ 1,200 nm(2)/s. By contrast, our Molecular Dynamics simulations show that diffusion of an isolated protein under force occurs at D(eff) ~ 10(8) nm(2)/s. This discrepancy is attributed to the drag force caused by the tethering object. From the physiological time scales of tissue elasticity, we calculate that tethered elastic proteins equilibrate in vivo with D(eff) ~ 10(4)-10(6) nm(2)/s which is two to four orders magnitude smaller than the values measured for untethered proteins in bulk.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Elasticity, structure, and relaxation of extended proteins under force.Proc Natl Acad Sci U S A. 2013 Mar 5;110(10):3847-52. doi: 10.1073/pnas.1300596110. Epub 2013 Feb 13. Proc Natl Acad Sci U S A. 2013. PMID: 23407163 Free PMC article.
-
Squeezing protein shells: how continuum elastic models, molecular dynamics simulations, and experiments coalesce at the nanoscale.Biophys J. 2010 Aug 9;99(4):1175-81. doi: 10.1016/j.bpj.2010.05.033. Biophys J. 2010. PMID: 20713001 Free PMC article.
-
Improved resolution of tertiary structure elasticity in muscle protein.Biophys J. 2011 Feb 16;100(4):L22-4. doi: 10.1016/j.bpj.2011.01.019. Biophys J. 2011. PMID: 21320420 Free PMC article.
-
AFM review study on pox viruses and living cells.Biophys J. 1997 Oct;73(4):2183-94. doi: 10.1016/S0006-3495(97)78250-X. Biophys J. 1997. PMID: 9336215 Free PMC article. Review.
-
Entropic elastic processes in protein mechanisms. II. Simple (passive) and coupled (active) development of elastic forces.J Protein Chem. 1988 Apr;7(2):81-114. doi: 10.1007/BF01025240. J Protein Chem. 1988. PMID: 3076450 Review.
Cited by
-
The elastic free energy of a tandem modular protein under force.Biochem Biophys Res Commun. 2015 May 1;460(2):434-8. doi: 10.1016/j.bbrc.2015.03.051. Epub 2015 Mar 18. Biochem Biophys Res Commun. 2015. PMID: 25796331 Free PMC article.
-
Next Generation Methods for Single-Molecule Force Spectroscopy on Polyproteins and Receptor-Ligand Complexes.Front Mol Biosci. 2020 May 19;7:85. doi: 10.3389/fmolb.2020.00085. eCollection 2020. Front Mol Biosci. 2020. PMID: 32509800 Free PMC article. Review.
-
On artifacts in single-molecule force spectroscopy.Proc Natl Acad Sci U S A. 2015 Nov 17;112(46):14248-53. doi: 10.1073/pnas.1519633112. Epub 2015 Nov 4. Proc Natl Acad Sci U S A. 2015. PMID: 26540730 Free PMC article.
-
Force dependency of biochemical reactions measured by single-molecule force-clamp spectroscopy.Nat Protoc. 2013;8(7):1261-76. doi: 10.1038/nprot.2013.056. Epub 2013 Jun 6. Nat Protoc. 2013. PMID: 23744288 Free PMC article.
-
Analytical Approaches for Deriving Friction Coefficients for Selected α-Helical Peptides Based Entirely on Molecular Dynamics Simulations.J Phys Chem B. 2022 Nov 10;126(44):8901-8912. doi: 10.1021/acs.jpcb.2c03076. Epub 2022 Oct 27. J Phys Chem B. 2022. PMID: 36300354 Free PMC article.
References
-
- Linke WA. Sense and stretchability: The role of titin and titin-associated proteins in myocardial stress-sensing and mechanical dysfunction. Cardiovasc Res. 2008;77:637–648. - PubMed
-
- Li HB, et al. Reverse engineering of the giant muscle protein titin. Nature. 2002;418:998–1002. - PubMed
-
- Greenewalt CH. The wings of insects and birds as mechanical oscillators. Proc Am Philos Soc. 1960;104:605–611.
-
- Deban SM, Richardson JC. Cold-blooded snipers: Thermal independence of ballistic tongue projection in the salamander hydromantes platycephalus. J Exp Zool A. 2011;315A:618–630. - PubMed
-
- Roberts TJ, Marsh RL, Weyand PG, Taylor CR. Muscular force in running turkeys: The economy of minimizing work. Science. 1997;275:1113–1115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
