

Energy balance modulates mouse skin tumor promotion through altered IGF-1R and EGFR crosstalk
- PMID: 22896210
- PMCID: PMC3482456
- DOI: 10.1158/1940-6207.CAPR-12-0234
Energy balance modulates mouse skin tumor promotion through altered IGF-1R and EGFR crosstalk
Abstract
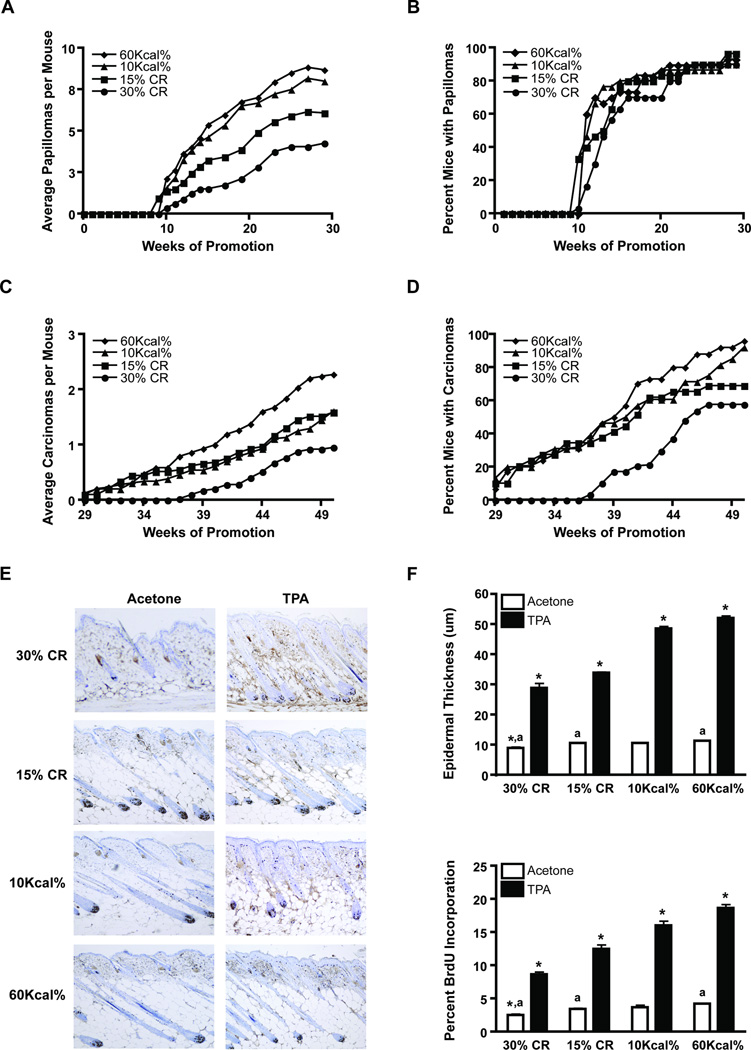
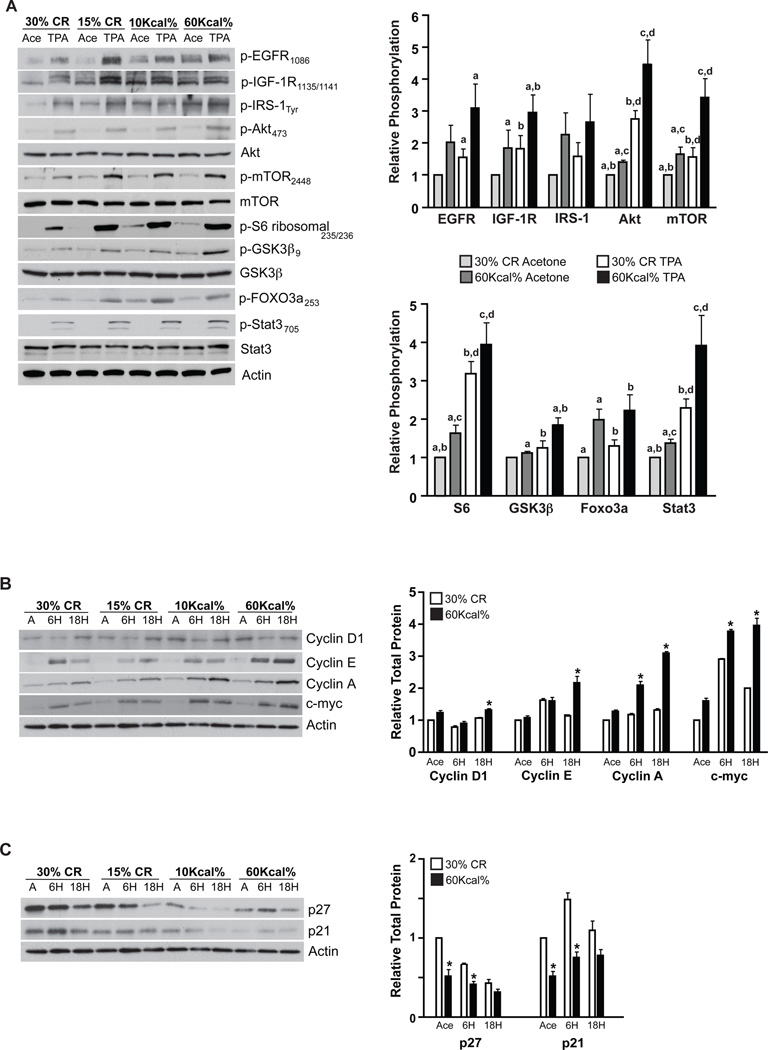
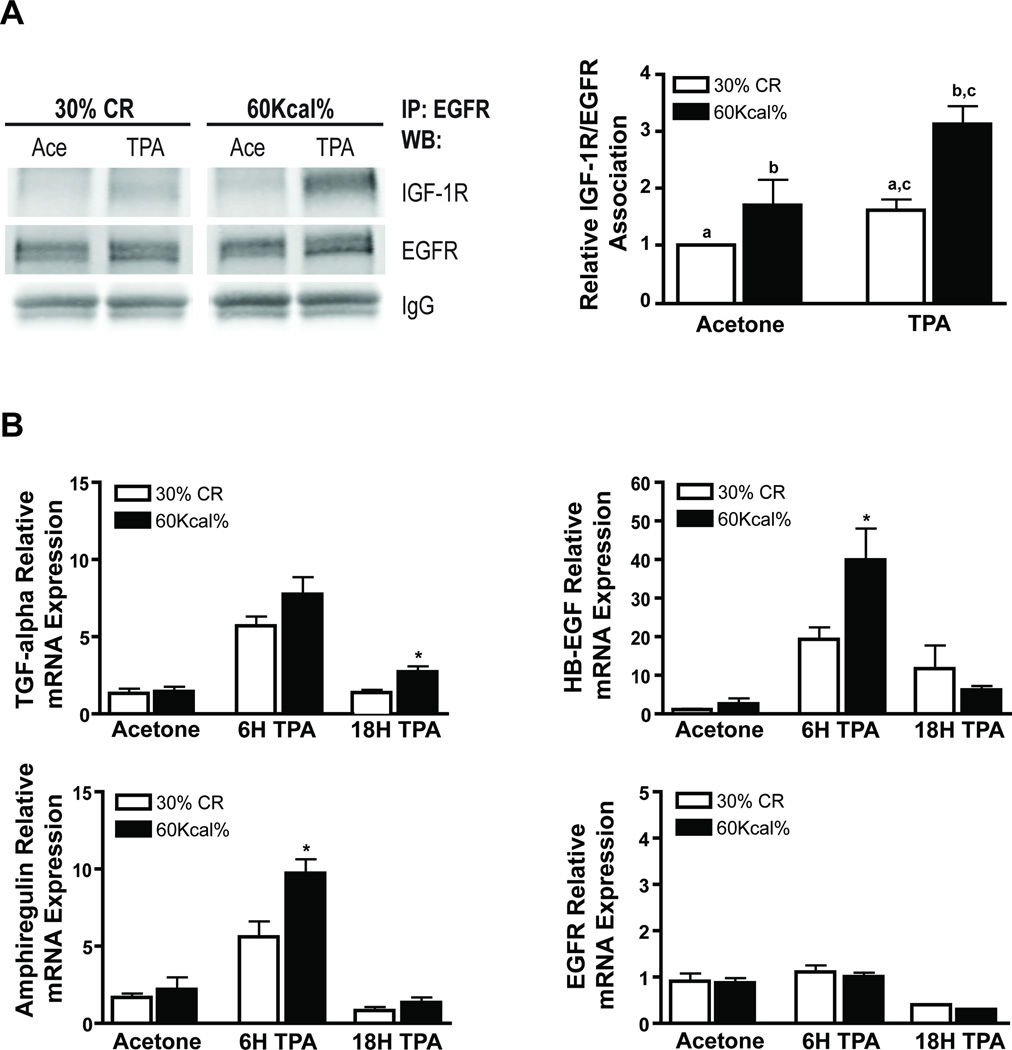
Obesity, an established risk factor for epithelial cancers, remains prevalent in the United States and many other countries. In contrast to positive energy balance states (overweight, obesity), calorie restriction (CR) has been shown to act as a universal inhibitor of tumorigenesis in multiple animal models of human cancer. Unfortunately, the mechanisms underlying the enhancing effects of obesity or the inhibitory effects of CR on cancer etiology remain elusive. Here, we evaluated the impact of dietary energy balance manipulation on epithelial carcinogenesis and identified several potential mechanisms that may account for the differential effects of obesity and CR on cancer. Obesity enhanced tumor promotion during epithelial carcinogenesis, in part, due to altered insulin-like growth factor-1 receptor (IGF-1R)/EGF receptor (EGFR) crosstalk and downstream signaling to effectors such as Akt/mTOR. Obesity-induced changes in cellular signaling subsequently led to altered levels of cell-cycle proteins that favored enhanced epidermal proliferation during tumor promotion. In contrast, CR reduced susceptibility to tumor promotion, attenuated IGF-1R/EGFR crosstalk and downstream signaling, and altered levels of cell-cycle proteins that favored reduced epidermal proliferation during tumor promotion. Collectively, these findings suggest potential targets for the prevention of epithelial cancers, as well as for reversal of obesity-mediated cancer development and progression. Cancer Prev Res; 5(10); 1236-46. ©2012 AACR.
Conflict of interest statement
There are no conflicts of interest.
Figures


References
-
- Patel AC, Nunez NP, Perkins SN, Barrett JC, Hursting SD. Effects of energy balance on cancer in genetically altered mice. J Nutr. 2004;134:3394S–3398S. - PubMed
-
- Calle EE, Kaaks R. Overweight, obesity and cancer: epidemiological evidence and proposed mechanisms. Nat Rev Cancer. 2004;4:579–591. - PubMed
-
- Yakar S, Nunez NP, Pennisi P, Brodt P, Sun H, Fallavollita L, et al. Increased Tumor Growth in Mice with Diet-Induced Obesity: Impact of Ovarian Hormones. Endocrinology. 2006 - PubMed
-
- Nunez NP, Perkins SN, Smith NC, Berrigan D, Berendes DM, Varticovski L, et al. Obesity accelerates mouse mammary tumor growth in the absence of ovarian hormones. Nutr Cancer. 2008;60:534–541. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
