

Secretomic analysis identifies alpha-1 antitrypsin (A1AT) as a required protein in cancer cell migration, invasion, and pericellular fibronectin assembly for facilitating lung colonization of lung adenocarcinoma cells
- PMID: 22896658
- PMCID: PMC3494203
- DOI: 10.1074/mcp.M112.017384
Secretomic analysis identifies alpha-1 antitrypsin (A1AT) as a required protein in cancer cell migration, invasion, and pericellular fibronectin assembly for facilitating lung colonization of lung adenocarcinoma cells
Abstract
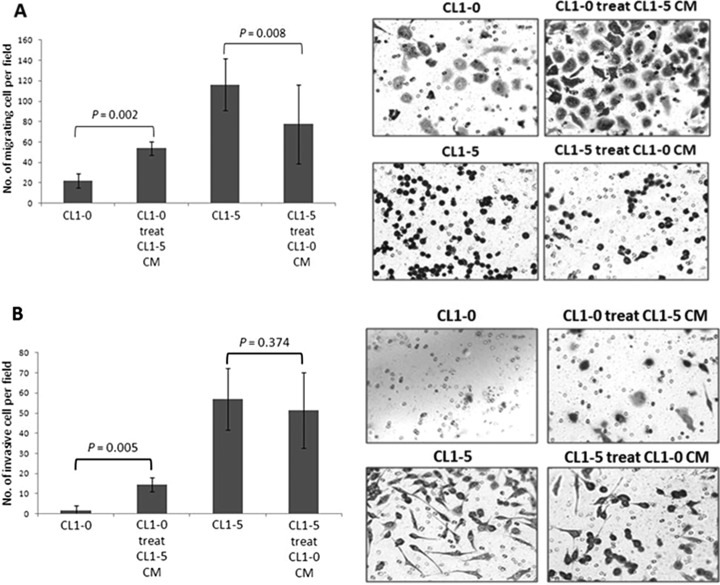
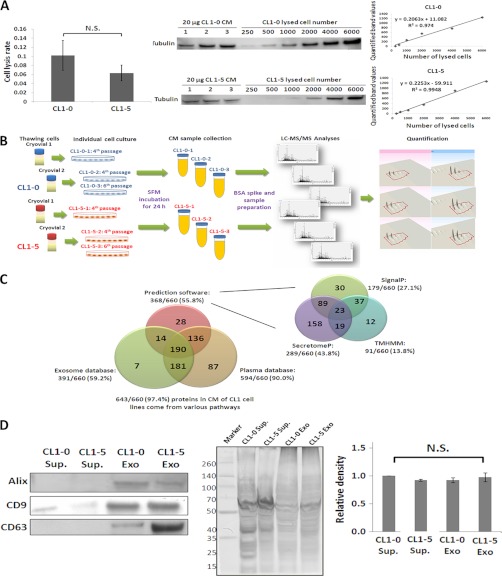
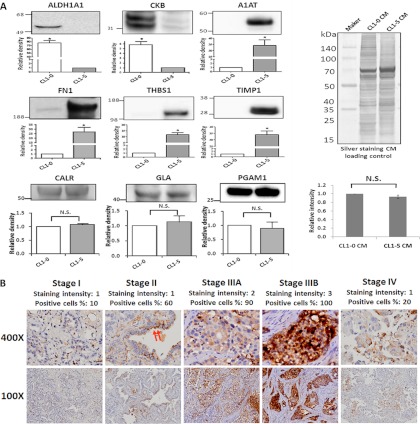
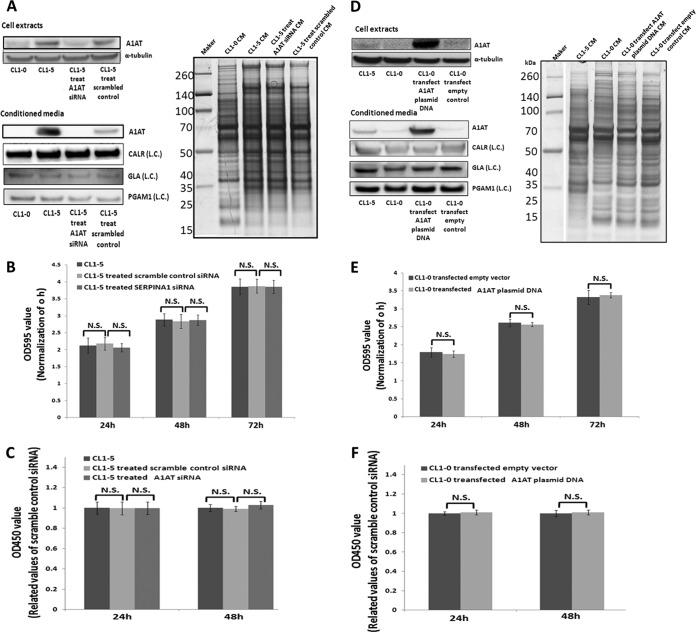
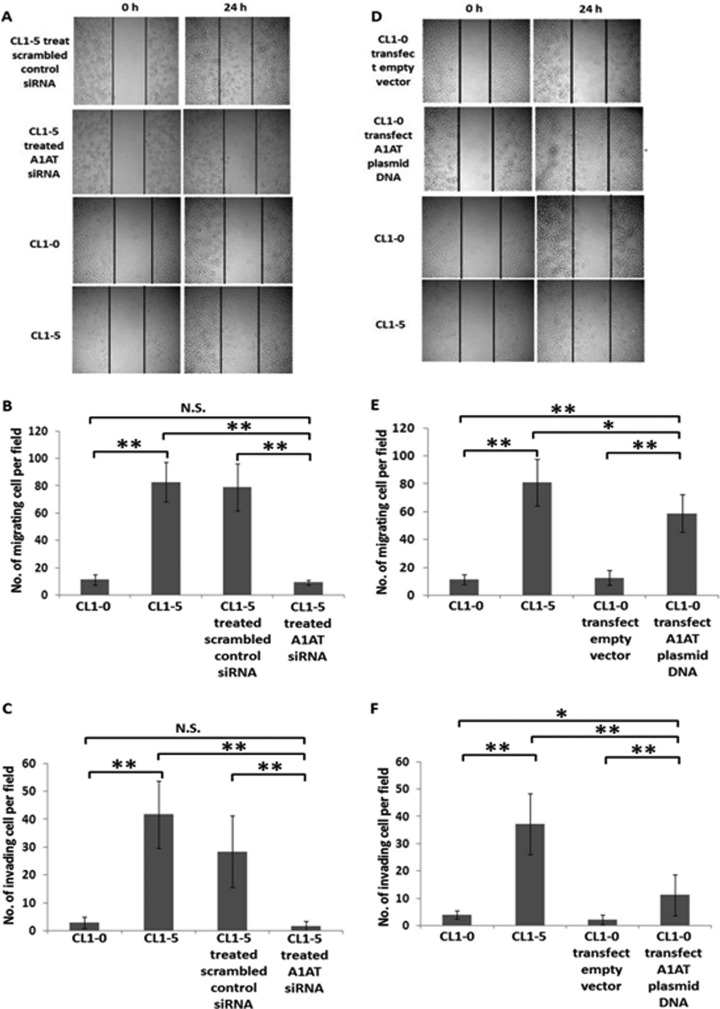
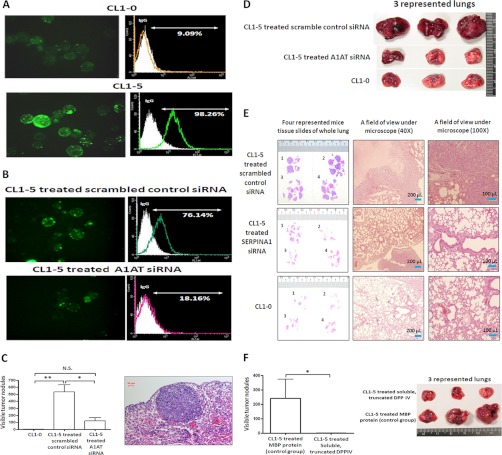
Metastasis is a major obstacle that must be overcome for the successful treatment of lung cancer. Proteins secreted by cancer cells may facilitate the progression of metastasis, particularly within the phases of migration and invasion. To discover metastasis-promoting secretory proteins within cancer cells, we used the label-free quantitative proteomics approach and compared the secretomes from the lung adenocarcinoma cell lines CL1-0 and CL1-5, which exhibit low and high metastatic properties, respectively. By employing quantitative analyses, we identified 660 proteins, 68 of which were considered to be expressed at different levels between the two cell lines. High levels of A1AT were secreted by CL1-5, and the roles of A1AT in the influence of lung adenocarcinoma metastasis were investigated. Molecular and pathological confirmation demonstrated that altered expression of A1AT correlates with the metastatic potential of lung adenocarcinoma. The migration and invasion properties of CL1-5 cells were significantly diminished by reducing the expression and secretion of their A1AT proteins. Conversely, the migration and invasion properties of CL1-0 cells were significantly increased through the overexpression and secretion of A1AT proteins. Furthermore, the assembly levels of the metastasis-promoting pericellular fibronectin (FN1), which facilitates colonization of lung capillary endothelia by adhering to the cell surface receptor dipeptidyl peptidase IV (DPP IV), were higher on the surfaces of suspended CL1-5 cells than on those of the CL1-0 cells. This discovery reflects previous findings in breast cancer. In line with this finding, FN1 assembly and the lung colonization of suspended CL1-5 cells were inhibited when endogenous A1AT protein was knocked down using siRNA. The major thrust of this study is to demonstrate the effects of coupling the label-free proteomics strategy with the secretomes of cancer cells that differentially exhibit invasive and metastatic properties. This provides a new opportunity for the effective identification of metastasis-associated proteins that are secreted by cancer cells and promote experimental metastasis.
Figures
 ), generic kinase (
), generic kinase ( ), metalloprotease (
), metalloprotease ( ), generic protease (
), generic protease ( ), generic binding protein (
), generic binding protein ( ), lipid phosphatase (
), lipid phosphatase ( ), generic receptor (
), generic receptor ( ), receptor ligand (
), receptor ligand ( ), positive effect (
), positive effect ( ), negative effect (
), negative effect ( ), unspecified effect (
), unspecified effect ( ), the proposed possible pathway (
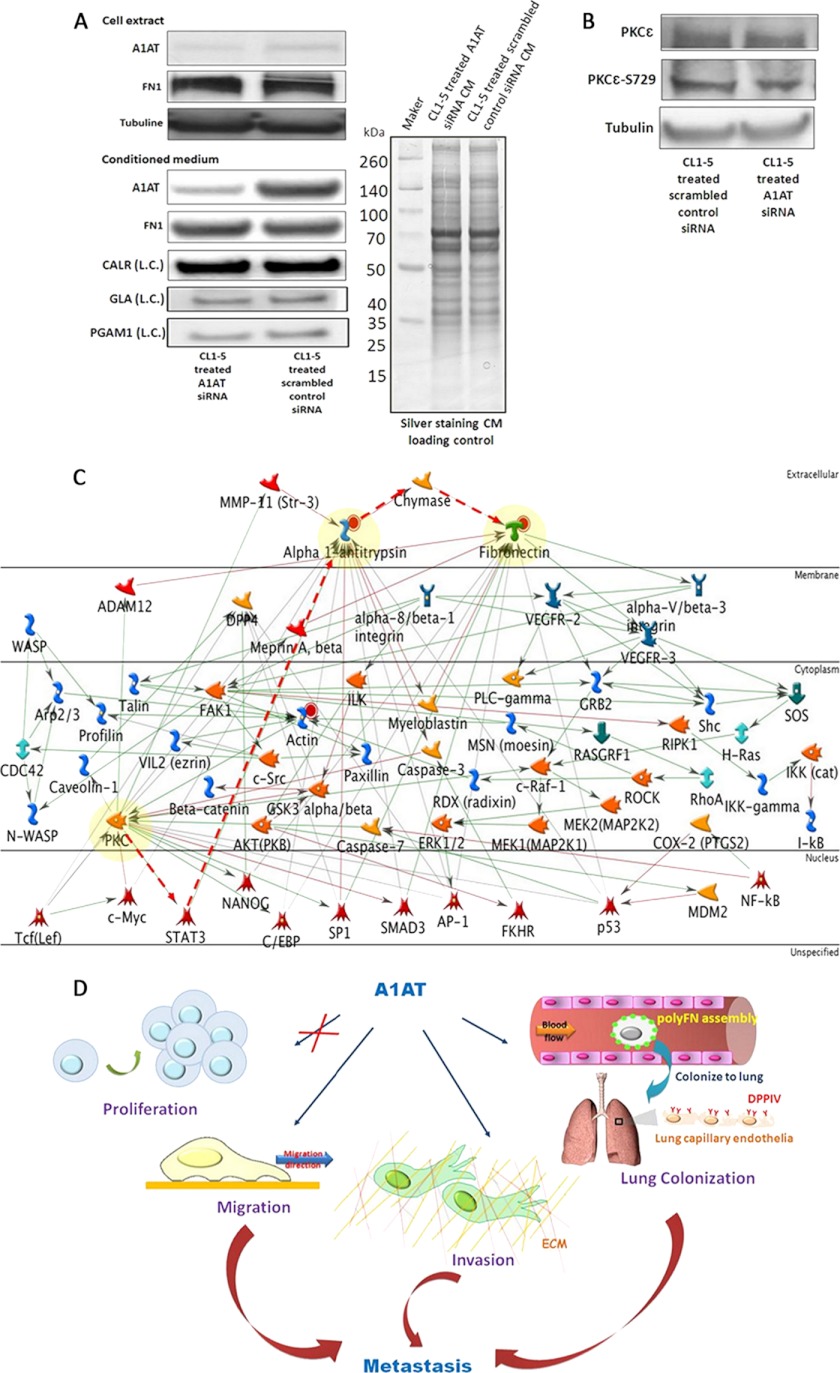
), the proposed possible pathway ( ). D, This scheme summarizes the functions in which A1AT is involved when facilitating lung adenocarcinoma experimental metastasis. A1AT mediates migration and invasion abilities but not the proliferation of cancer cells. Additionally, it regulates polyFN assembly on lung adenocarcinoma cell surfaces to colonize on the lung via adhering to DPP IV on lung capillary endothelia.
). D, This scheme summarizes the functions in which A1AT is involved when facilitating lung adenocarcinoma experimental metastasis. A1AT mediates migration and invasion abilities but not the proliferation of cancer cells. Additionally, it regulates polyFN assembly on lung adenocarcinoma cell surfaces to colonize on the lung via adhering to DPP IV on lung capillary endothelia.References
-
- Hanahan D., Weinberg R. A. (2000) The hallmarks of cancer. Cell. 100, 57–70 - PubMed
-
- D'Addario G., Früh M., Reck M., Baumann P., Klepetko W., Felip E., and ESMO Guidelines Working Group (2010) Metastatic non-small-cell lung cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 21, 116–119 - PubMed
-
- Woodhouse E. C., Chuaqui R. F., Liotta L. A. (1997) General mechanisms of metastasis. Cancer 80, 1529–1537 - PubMed
-
- Box G. M., Eccles S. A. (2011) Simple experimental and spontaneous metastasis assays in mice. Methods Mol Biol. 769, 311–329 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
