

Genetically engineered fluorescent voltage reporters
- PMID: 22896802
- PMCID: PMC3419450
- DOI: 10.1021/cn300041b
Genetically engineered fluorescent voltage reporters
Abstract
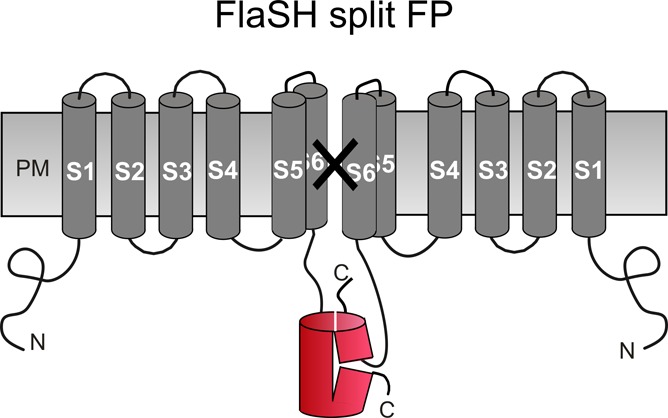
Fluorescent membrane voltage indicators that enable optical imaging of neuronal circuit operations in the living mammalian brain are powerful tools for biology and particularly neuroscience. Classical voltage-sensitive dyes, typically low molecular-weight organic compounds, have been in widespread use for decades but are limited by issues related to optical noise, the lack of generally applicable procedures that enable staining of specific cell populations, and difficulties in performing imaging experiments over days and weeks. Genetically encoded voltage indicators (GEVIs) represent a newer alternative that overcomes several of the limitations inherent to classical voltage-sensitive dyes. We critically review the fundamental concepts of this approach, the variety of available probes and their state of development.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
