

Synergy between PI3K signaling and MYC in Burkitt lymphomagenesis
- PMID: 22897848
- PMCID: PMC3432451
- DOI: 10.1016/j.ccr.2012.06.012
Synergy between PI3K signaling and MYC in Burkitt lymphomagenesis
Abstract
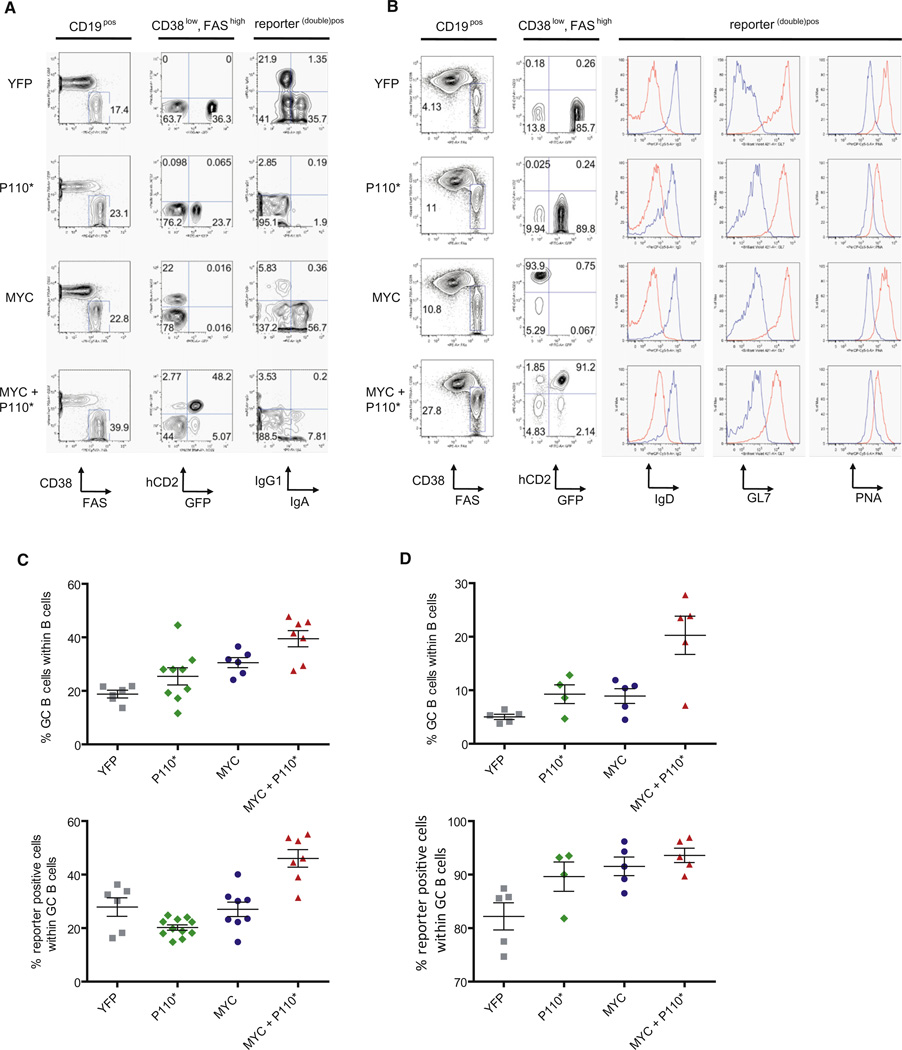
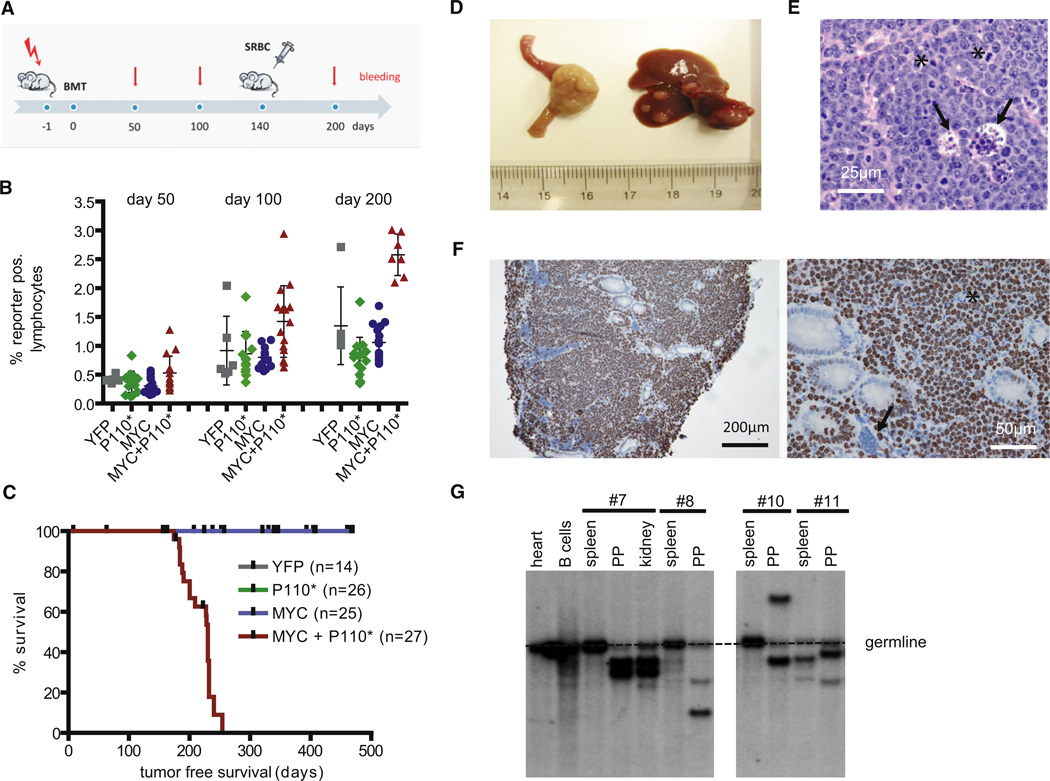
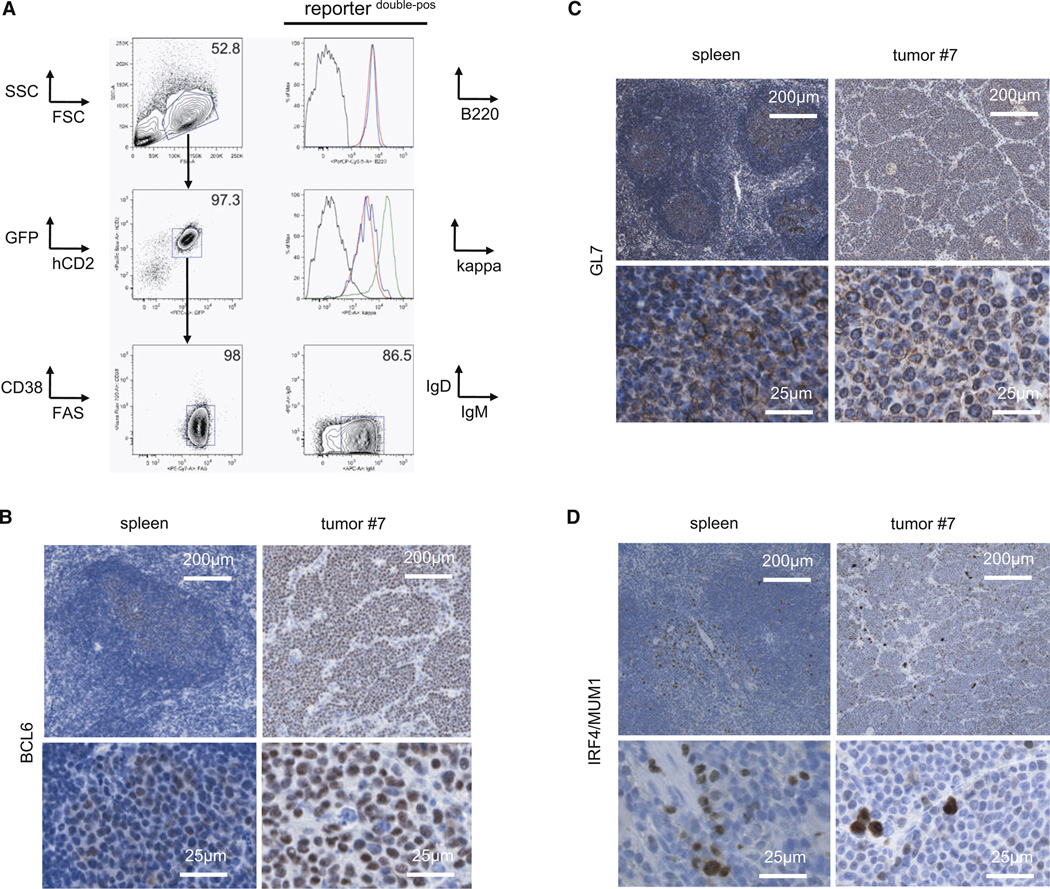
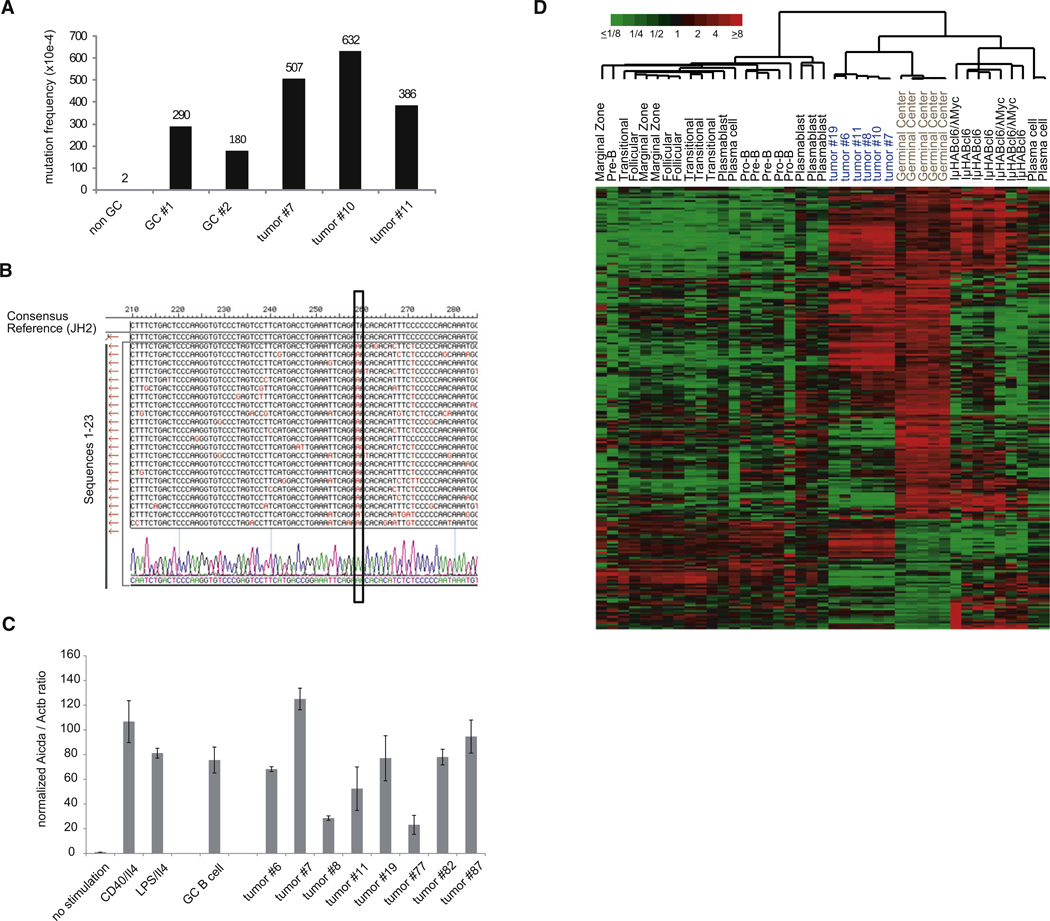
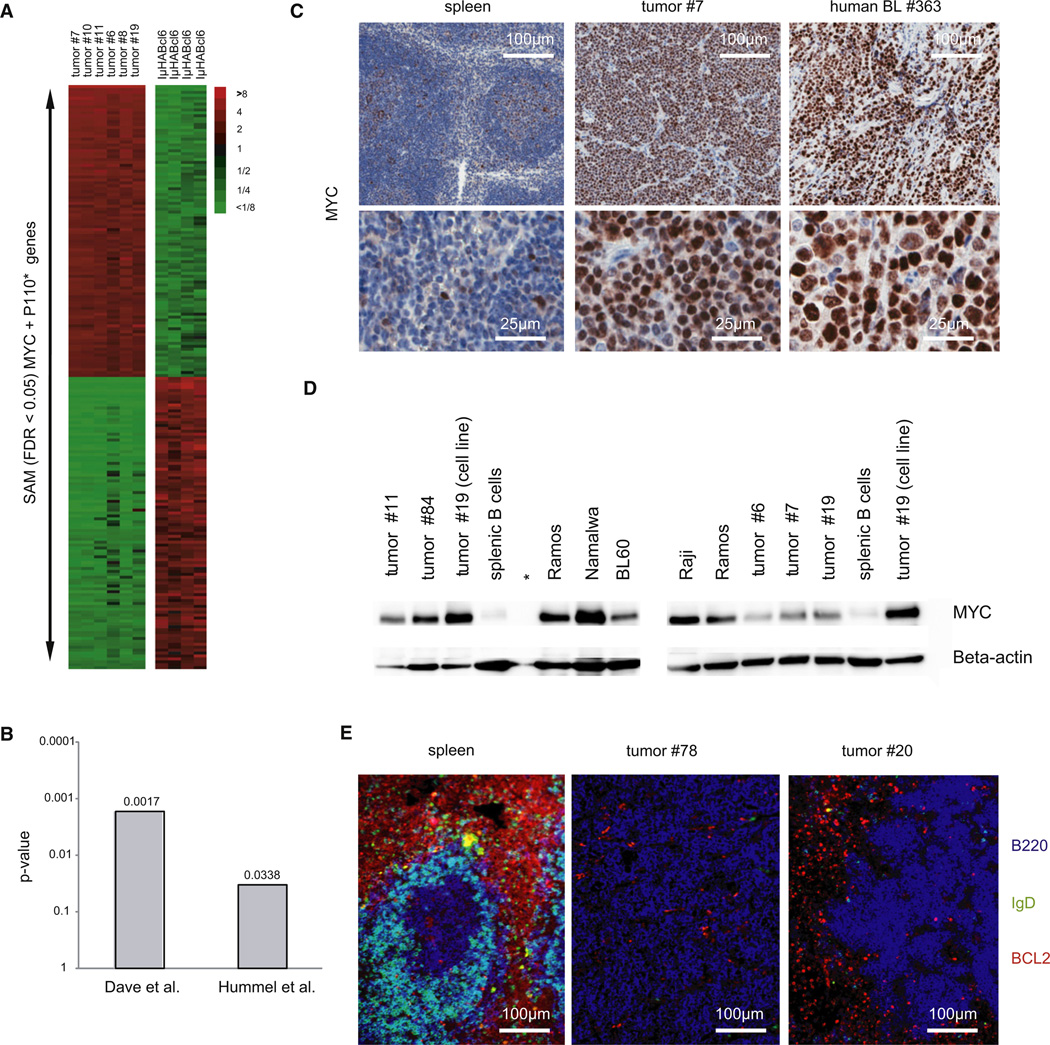
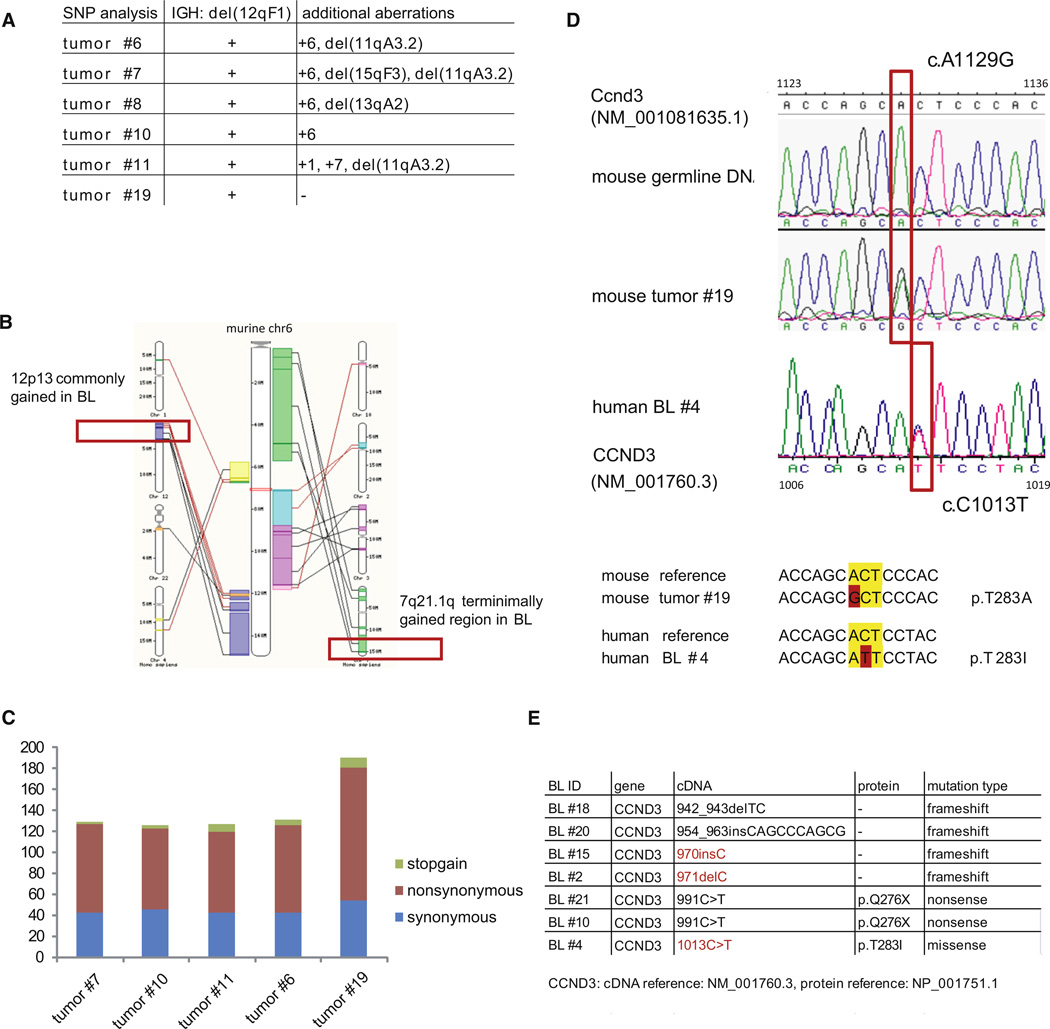
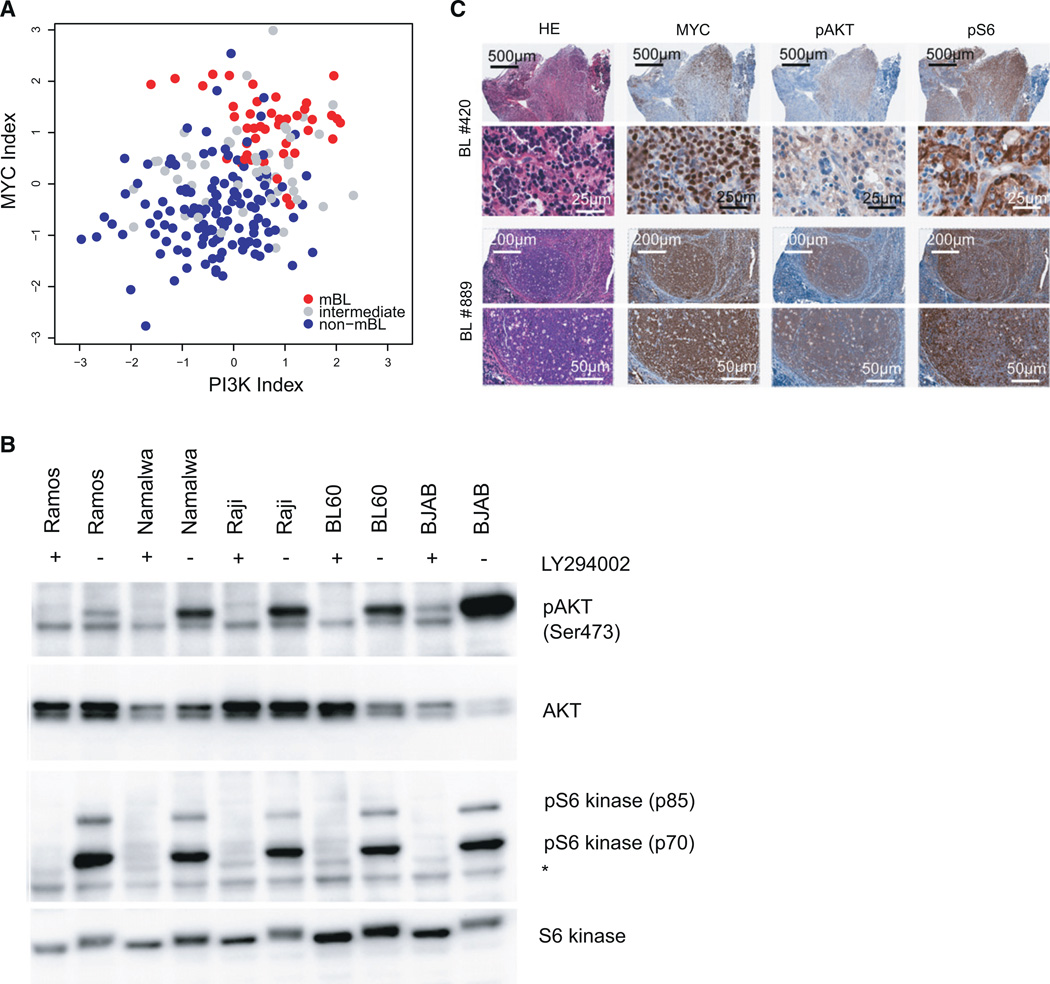
In Burkitt lymphoma (BL), a germinal center B-cell-derived tumor, the pro-apoptotic properties of c-MYC must be counterbalanced. Predicting that survival signals would be delivered by phosphoinositide-3-kinase (PI3K), a major survival determinant in mature B cells, we indeed found that combining constitutive c-MYC expression and PI3K activity in germinal center B cells of the mouse led to BL-like tumors, which fully phenocopy human BL with regard to histology, surface and other markers, and gene expression profile. The tumors also accumulate tertiary mutational events, some of which are recurrent in the human disease. These results and our finding of recurrent PI3K pathway activation in human BL indicate that deregulated c-MYC and PI3K activity cooperate in BL pathogenesis.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Burkitt lymphoma: much more than MYC.Cancer Cell. 2012 Aug 14;22(2):141-2. doi: 10.1016/j.ccr.2012.07.018. Cancer Cell. 2012. PMID: 22897844
-
Targeted therapies: New targets in Burkitt lymphoma?Nat Rev Clin Oncol. 2012 Oct;9(10):551. doi: 10.1038/nrclinonc.2012.153. Epub 2012 Aug 28. Nat Rev Clin Oncol. 2012. PMID: 22926023 No abstract available.
References
-
- Adams JM, Cory S. Myc oncogene activation in B and T lymphoid tumours. Proc. R. Soc. Lond. B Biol. Sci. 1985;226:59–72. - PubMed
-
- Alizadeh AA, Eisen MB, Davis RE, Ma C, Lossos IS, Rosenwald A, Boldrick JC, Sabet H, Tran T, Yu X, et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature. 2000;403:503–511. - PubMed
-
- Bengtsson H, Irizarry R, Carvalho B, Speed TP. Estimation and assessment of raw copy numbers at the single locus level. Bioinformatics. 2008;24:759–767. - PubMed
-
- Bhatia KG, Gutiérrez MI, Huppi K, Siwarski D, Magrath IT. The pattern of p53 mutations in Burkitt’s lymphoma differs from that of solid tumors. Cancer Res. 1992;52:4273–4276. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
