

doi: 10.1016/j.ccr.2012.06.013.
Small GTPase R-Ras regulates integrity and functionality of tumor blood vessels
Affiliations
- PMID: 22897853
- PMCID: PMC3422514
- DOI: 10.1016/j.ccr.2012.06.013
Item in Clipboard
Small GTPase R-Ras regulates integrity and functionality of tumor blood vessels
Cancer Cell.
.
Abstract
We show that R-Ras, a small GTPase of the Ras family, is essential for the establishment of mature, functional blood vessels in tumors. The genetic disruption of R-Ras severely impaired the maturation processes of tumor vessels in mice. Conversely, the gain of function of R-Ras improved vessel structure and blood perfusion and blocked plasma leakage by enhanced endothelial barrier function and pericyte association with nascent blood vessels. Thus, R-Ras promotes normalization of the tumor vasculature. These findings identify R-Ras as a critical regulator of vessel integrity and function during tumor vascularization.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
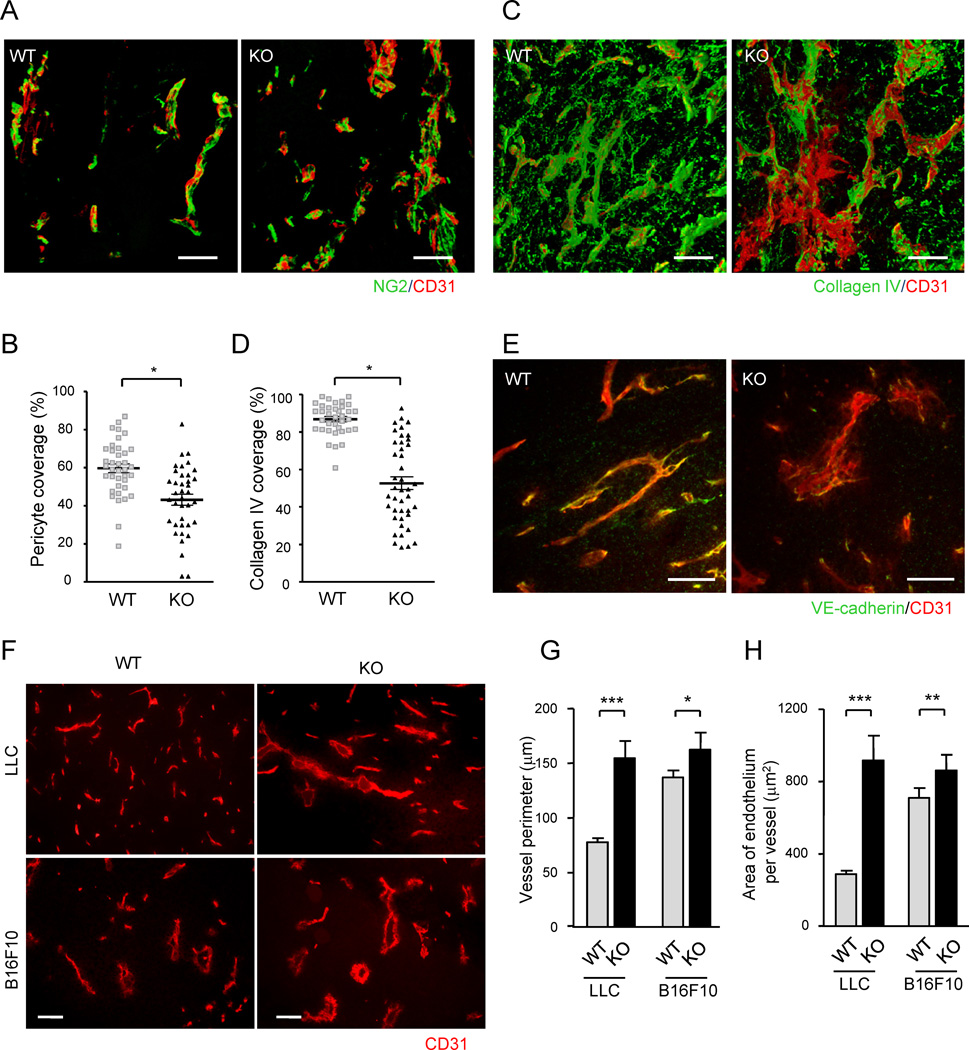
(A) Endothelium and associated pericytes were visualized by CD31 (red) and NG2 (green) immunofluorescence staining of B16F10 tumor implants. Representative images of wild type (WT) and R-Ras KO (KO) tumor vessels are shown. See also Video S1. (B) Pericyte coverage was quantified by calculating the % fraction of vessel surface (CD31) area that overlapped with NG2 staining in the 3-D image of the vessels to determine direct contact between the two cell types. *p = 0.001, ±SEM. (C and D) A 3-D analysis of collagen IV coverage. CD31, red; collagen IV, green. *p = 4×10−13 (E) Double staining of LLC tumors for CD31 (red) and VE-cadherin (green). (F) CD31-staining of LLC and B16F10 tumor sections. Scale bars, 100 µm (A, C, F), 50 µm (E). (G and H) The perimeter and thickness of the endothelium (assessed by average CD31+ area per vessel) of tumor vessels were increased in R-Ras KO mice, reflecting the augmented vessel deformations. *p = 0.01, **p < 0.001, ***p < 1×10−4
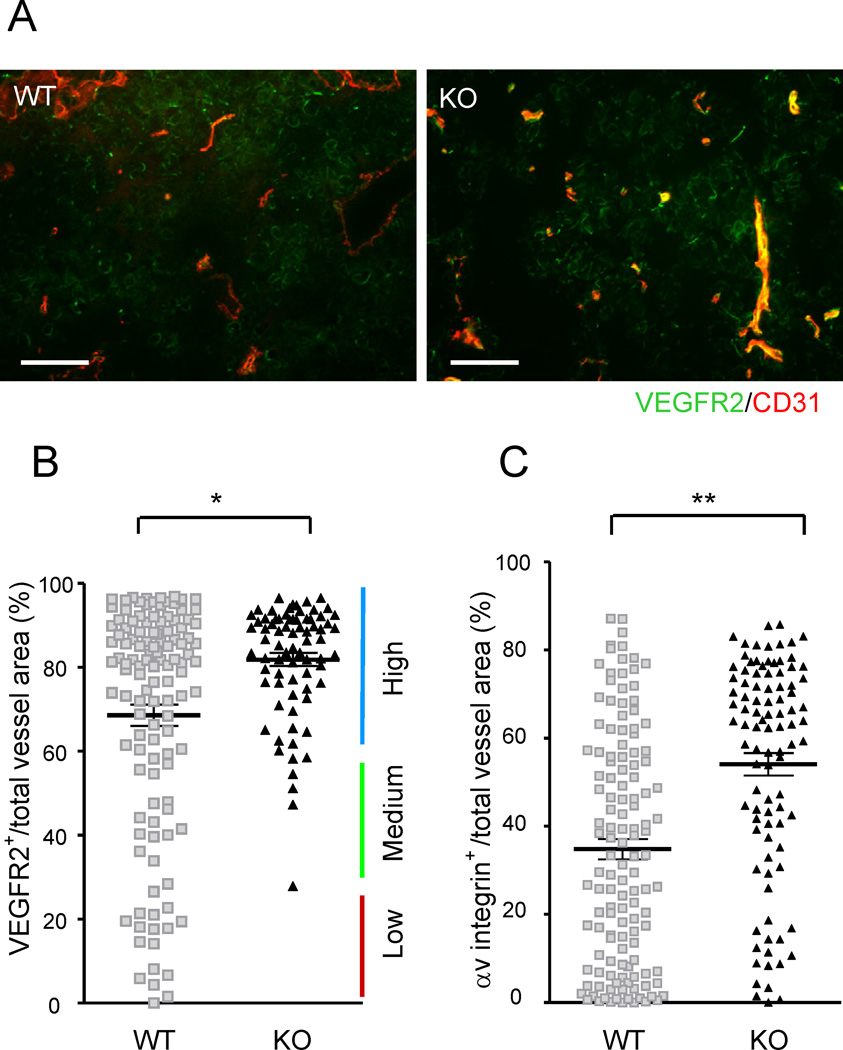
(A) Immunostaining of B16F10 tumor implants for CD31 (red) and VEGFR2 (green). Yellow, double-stained area. (B) The VEGFR2 expression status of the tumor vasculature was assessed by the fraction of VEGFR2-expressing vessel area (VEGFR2+CD31+ double-positive) as a percentage of the total vessel area (total CD31). Each dot represents the average of several vessels found in a micrograph. Fifteen micrographs were obtained randomly from multiple tumor sections. Eight wild type and five R-Ras KO mice were examined. *p = 1×10−5, ±SEM. (C) A similar analysis with αv integrin staining as a marker for angiogenic vessels. Scale bar, 100 µm. **p = 2×10−11. See also Figure S1.
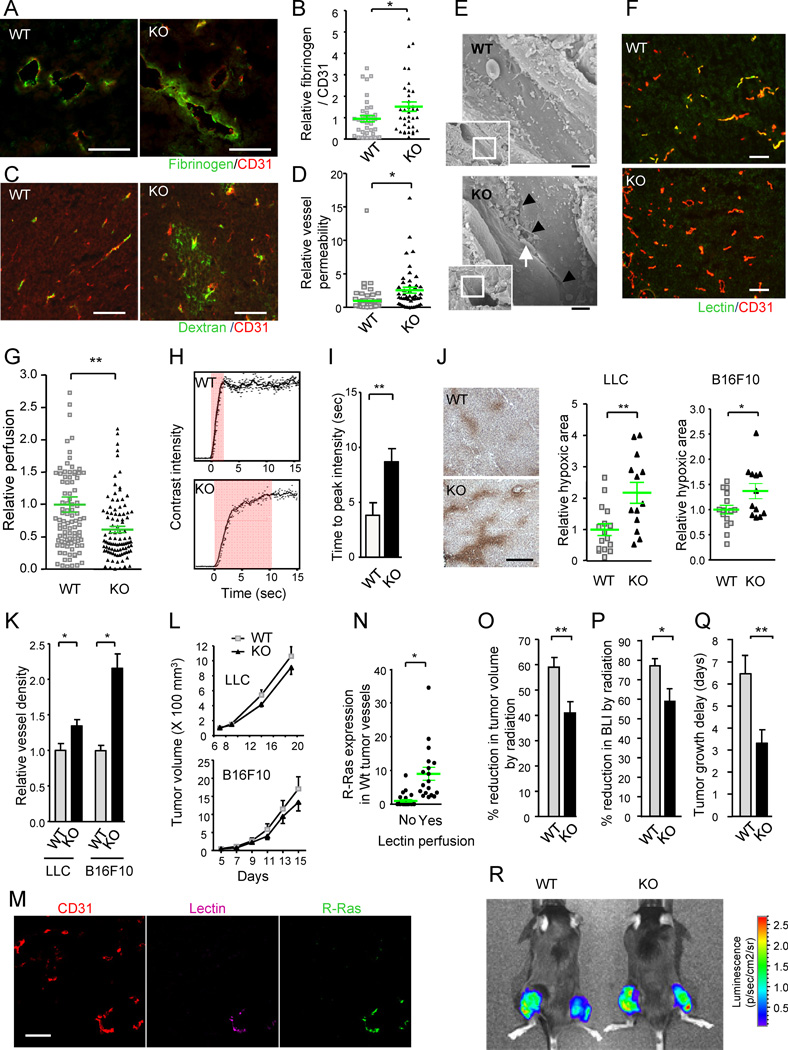
(A–D) Analyses of vessel leakiness. Fibrinogen and CD31 staining for plasma leakage in B16F10 tumors (A). To quantify the level of plasma leakage and standardize it for tumor vessel area, the ratio of fibrinogen (fibrin)-stained area to CD31 area was determined and is presented as relative values (B). *p = 0.035, ±SEM. Dextran-FITC was injected i.v. into mice bearing LLC tumors (C). The fluorescence intensity of extravasated FITC in the tumors was standardized for the perfused vessel counts to assess vessel permeability (D). *p = 0.002 (E) Ultrastructure of tumor vessels by scanning electron microscopy. The R-Ras KO tumor endothelium shows significant special gaps at the cell-cell junctions (arrowheads). Arrow, a red blood cell extravasating between the cellular gap. Scale bar, 2 µm. (F and G) Blood perfusion efficiency. Lectin perfusion and CD31 staining of B16F10 tumor sections (F). Yellow color indicates double-stained vessel area. Percent lectin+CD31+ double-positive area/total CD31+ area was determined to assess perfusion efficiency of tumor vasculature (G). **p = 2×10−4 (H and I) Study of tumor vessel perfusion by contrast-enhanced ultrasound imaging. Examples of raw contrast kinetics acquired after bolus i.v. injection of microbubble contrast agent (H). Delayed time-to-peak-intensity in KO mice indicating poor perfusion efficiency (I). **p = 0.02 (J) Analysis of tumor hypoxia. The ratio of the hypoxic area/total area of the tumor section was determined and presented as relative values. Representative anti-Hypoxyprobe staining of LLC tumors is shown. *p = 0.03, **p = 0.003 (K) Number of tumor vessels per unit area (vessel density) is presented relative to the control group. *p < 0.05 (L) Growth of LLC and B16F10 tumors. (M and N) Analysis of R-Ras expression in wild type vessels. Sections of lectin-perfused tumors were stained for CD31, lectin, and R-Ras (M). The tumor vessels were classified as lectin-perfused and non-perfused vessels, and the level of R-Ras expression was determined for individual vessels in each group (N). *p = 7×10−4
(O–R) Effect of radiotherapy. (O) The percent reduction in the tumor volume (as compared with untreated tumors) was determined at 7 days after a local 12 Gy irradiation of LLC tumors (O). **p < 0.01 (P) Percent inhibition of tumor growth was also assessed by bioluminescence tumor imaging at day 7. BLI, bioluminescence intensity. *p = 0.02 (Q) Tumor growth delay as defined by delay in tumor volume quadrupling time as a result of irradiation treatment. The average delay time (in days) is presented. **p < 0.01 (R) Bioluminescence imaging of Luc2-transfected LLC tumors. Tumors on the right thigh received 12Gy irradiation whereas tumors on the left did not. WT, wild type; KO, R-Ras KO mice. Scale bars, 100 µm (A, C, F, M), 200 µm (J). See also Figure S2 and Table S1.
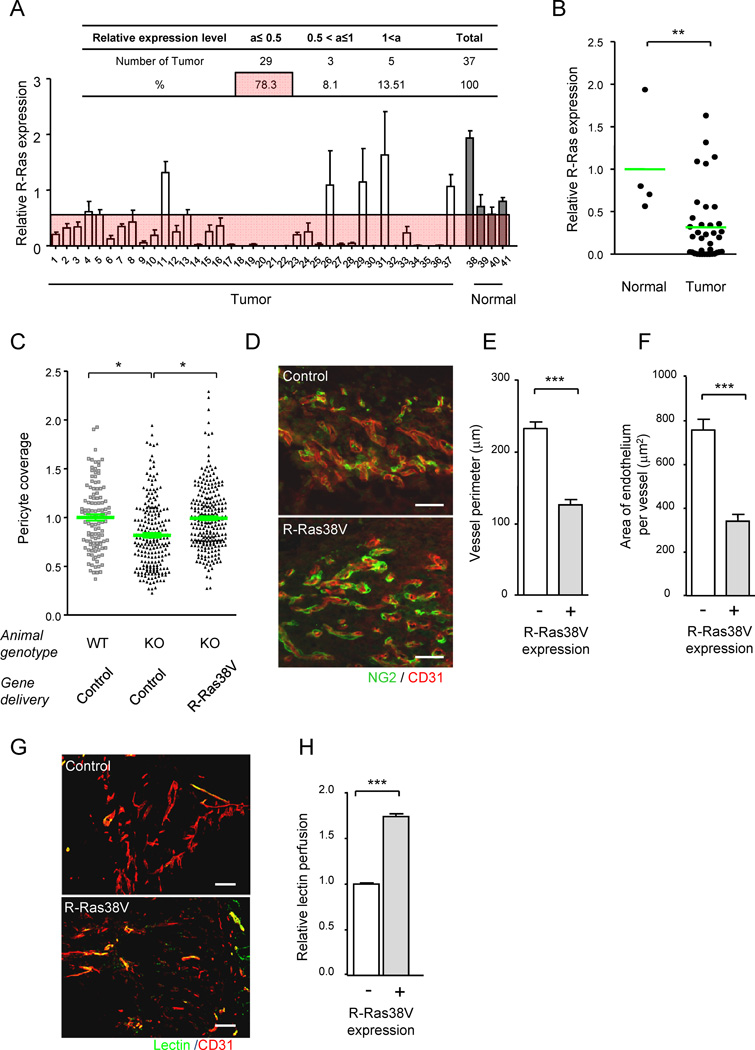
(A and B) R-Ras is downregulated in human tumor vasculature. The level of R-Ras expression in tumor vessels was analyzed in a human breast cancer tissue array by immunostaining for R-Ras and CD31 (A). Dot plot representation of A (B). **p = 0.004. See also Table S2. (C–H) R-Ras signaling improves structure and function of VEGF-induced vessels. VEGF-induced angiogenic vessels developed in Matrigel plugs were transduced in vivo with R-Ras38V or control lentivirus. Six days later, pericyte association with the endothelium was determined (C) by NG2 and CD31 immunofluorescence (D). The data is presented as relative values. (E–H) The vessels in the R-Ras38V virus-infected plugs were classified as R-Ras-positive (successfully transduced) or negative (non-transduced) vessels. The in vivo R-Ras transduction reduced the vessel perimeter (reduced vessel dilation) in R-Ras KO mice (E). The thickness of the endothelium (assessed by average CD31+ area per vessel) was also reduced in the KO mice by R-Ras transduction (F). The histological sections of Matrigel implants in R-Ras KO mice, which received control or R-Ras38V virus, were stained for perfused lectin, and CD31 to determine the blood perfusion of the vessels (G). Blood perfusion efficiency was compared between the R-Ras-transduced and non-transduced vessels (H). *p < 0.05, ***p < 1×10−3, ±SEM. Scale bars, 100 µm. See also Figure S3.
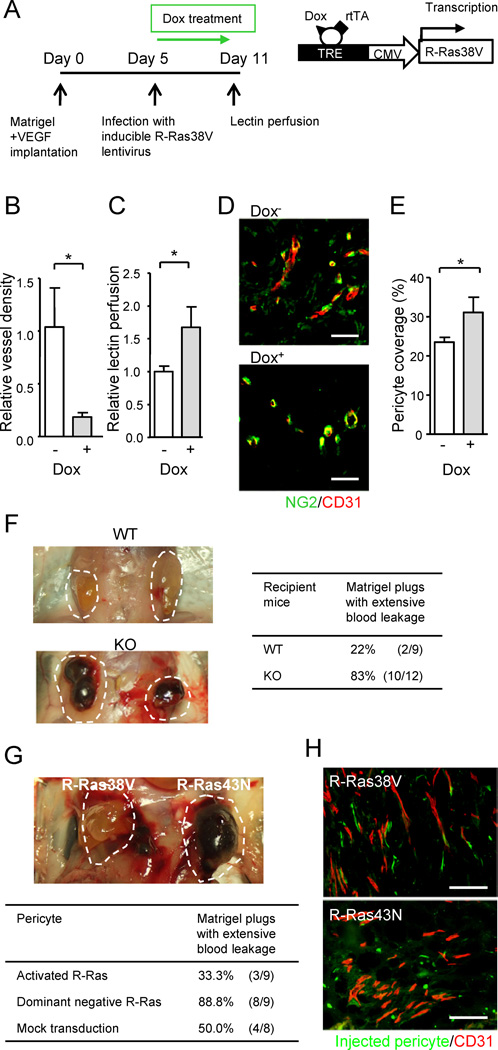
(A–E) The effect of endothelial specific R-Ras. A timetable of Matrigel implantation in Tie2-rtTA transgenic mice, lentivirus infection, R-Ras38V induction by doxycycline (Dox), and lectin i.v. perfusion is shown (A). Vessel density in the Matrigel plugs at Day 11 (B). Relative perfusion efficiency of i.v. injected lectin at Day 11 (C). (D and E) Matrigel sections were stained for NG2 and CD31 (D), and pericyte coverage of the vessels was determined by image analysis (E). (F) VEGF-induced Matrigel angiogenesis produces extensively ‘bloody’ plugs in R-Ras KO mice demonstrating enhanced leakiness of pathologically regenerating vessels in the absence of R-Ras. (G and H) The effect of pericyte specific R-Ras. Fluorescently labeled, ex-vivo-transduced pericytes were implanted with VEGF-containing Matrigel into the flank of R-Ras KO mice. Matrigel plugs were examined 7 days later (indicated by dashed lines) (G). Histological analysis of the plugs (H). The ex vivo-transduced, implanted pericytes were visualized by green fluorescence. Microvessels were stained with CD31 (red). Scale bars, 100 µm (H), 50 µm (D). *p < 0.05, ±SEM. See also Figure S4.
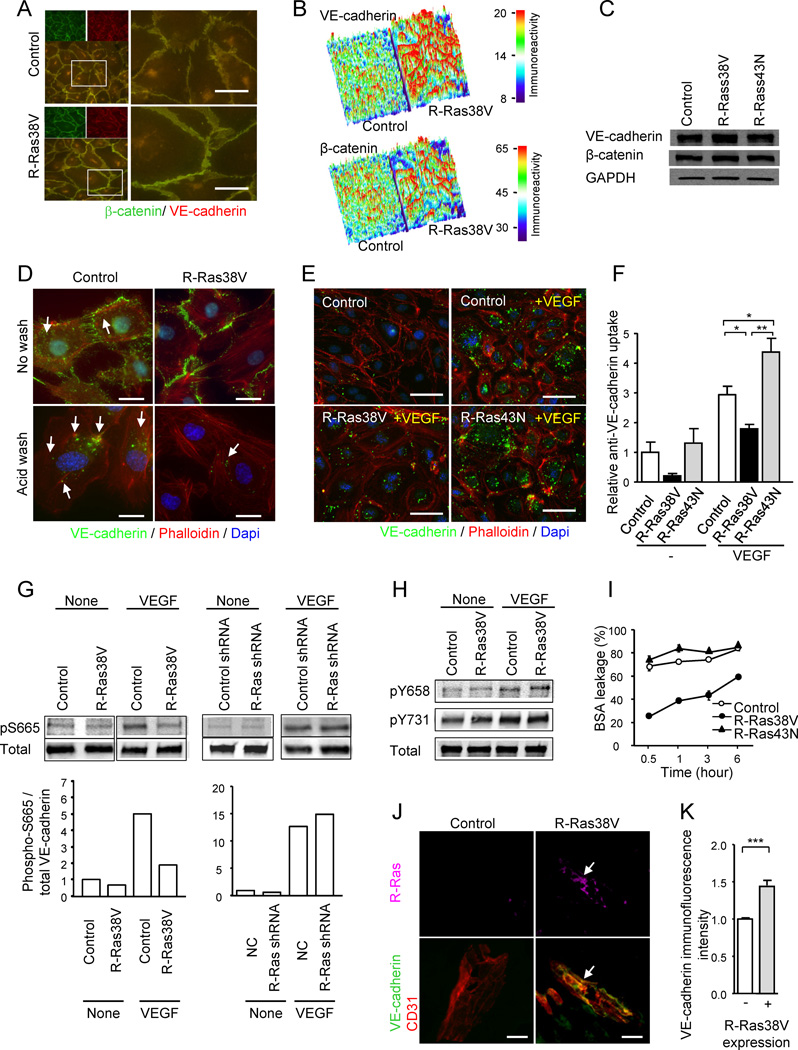
(A) R-Ras enhances adherens junction formation in EC monolayer. VE-cadherin (green) and β-catenin (red) staining of HUVEC monolayers transduced with R-Ras38V or control vector. Lower left panels and right panels are merged images of the double staining. Right panels show higher magnification at cell-cell junctions. (B) Surface plot of VE-cadherin and β-catenin immunofluorescence intensity shows marked accumulation of both proteins at the cell-cell interface upon R-Ras38V expression. The immunoreactivity is shown with arbitrary unit. (C) VE-cadherin and β-catenin immunoblotting of whole cell lysate. GAPDH is shown as a loading control. (D) Analysis of VEGF-induced VE-cadherin internalization. The monoclonal antibody (BV6) recognizes the extracellular domain of VE-cadherin (green). Following antibody incubation of the EC culture and stimulation with VEGF, the cell surface membrane-bound BV6 antibodies (upper panels) were removed by a mild acid wash (lower panels). The presence of internal acid-resistant vesicles stained with the BV6 antibody indicates the internalization of VE-cadherin. Phalloidin (actin), red; nucleus, blue; arrows, BV6-stained vesicles containing internalized VE-cadherin. (E) Dominant negative R-Ras43N enhances VE-cadherin internalization. (F) VE-cadherin internalization was quantified by the percentage of the cells showing internal acid-resistant vesicles. *p < 0.05, **p < 0.001, ±SEM. In the no-VEGF stimulation condition (−), statistical significance was not found between the control, R-Ras38V, and R-Ras43N. (G) Phosphorylation of VE-cadherin S665 was analyzed in the lysate of VEGF-stimulated ECs (100 ng/ml) using phospho-specific antibody. (H) Phosphorylation of VE-cadherin Y658 and Y731 residues. (I) The effect of R-Ras on the endothelial permeability was determined in HUVEC monolayer cultures. BSA leakage through the culture insert membrane without cells was set as 100%. (J) The in vivo transduction of R-Ras38V increased VE-cadherin immunoreactivity of angiogenic sprouts in Matrigel plugs implanted in R-Ras KO mice. Immunofluorescence showed accumulation of VE-cadherin at the endothelial cell-cell junctions upon R-Ras transgene expression (arrows). (K) Quantification of VE-cadherin immunoreactivity standardized with CD31 area was compared between R-Ras38V and mock transduced control plugs. ***p < 0.001. Scale bars, 20 µm (J), 25 µm (D), 50 µm (A, E). See also Figure S5.
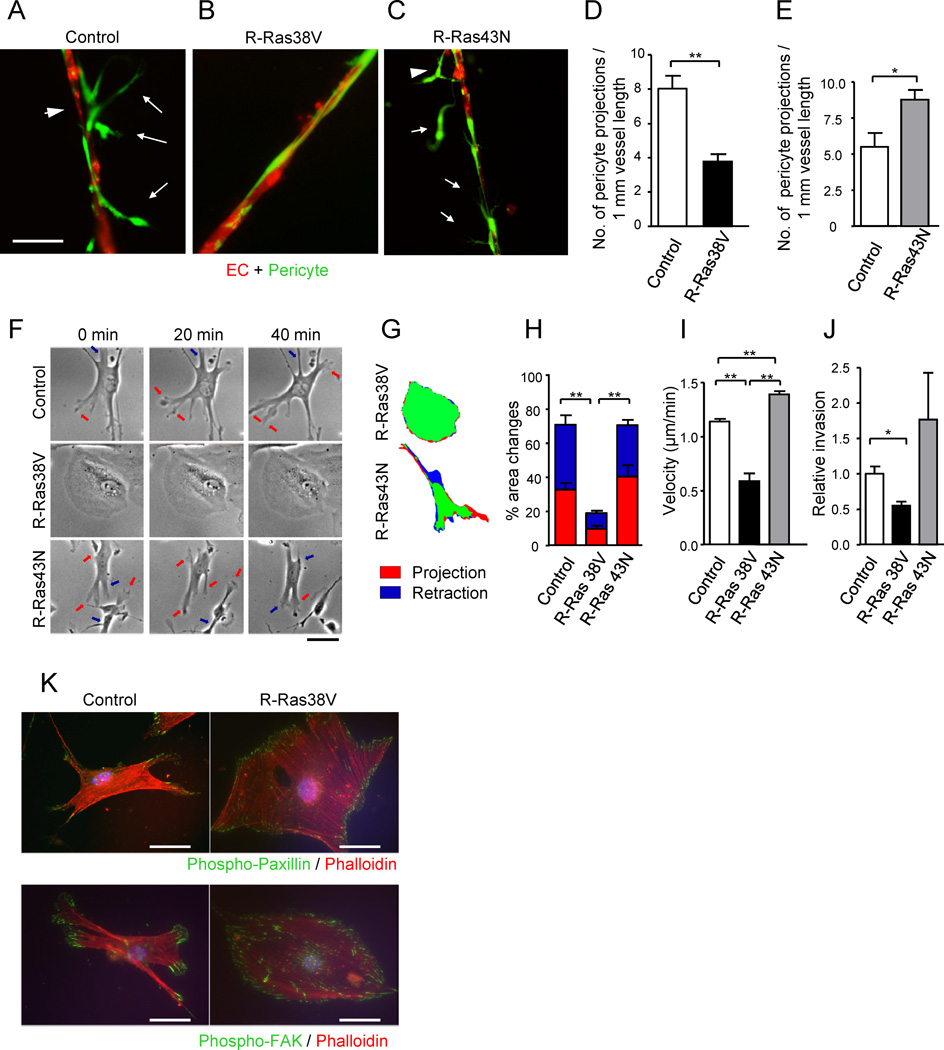
(A and B) Fluorescently labeled pericytes (green) were transduced with R-Ras38V or control vector and, at 3 hours post transduction, they were plated together with red-labeled ECs onto Matrigel at a 4:1 (EC : pericyte) ratio, cultured for 24 hours, and the resulting vessel-like structures were analyzed by fluorescent microscopy. In the control cocultures (A), pericytes were loosely attached to the vessel-like EC structures (arrowhead) with cytoplasmic processes (arrows). Upon R-Ras38V expression in pericytes, these abnormal characteristics were significantly diminished, and ECs and pericytes became tightly associated (B). (C) R-Ras43N expression in pericytes did not have this effect. (D and E) The number of pericyte projections from the reconstituted microvessels was significantly decreased by R-Ras38V (D) but increased by R-Ras43N expression (E). (F) Time-laps video microscopy of pericytes on the Matrigel-coated 2-D surface shows membrane projection/retraction activities of the pericytes. Arrows indicate dynamic membrane activities (orange, projections; blue, retractions). (G and H) The fractions of projecting (red) and retracting (blue) membrane area vs. total cell area (% area change) were determined between the time points 0 and 5 min. See also Figure S6. (I) The mean velocity of pericyte motility was calculated from 20 hour-long time-laps observations in 2-D culture. (J) Pericyte invasion through Matrigel-coated filter membrane. (K) Immunofluorescence of phospho-paxillin Y118 and phospho-FAK Y397 to highlight focal adhesions complexes (green) assembled throughout the cell perimeter upon R-Ras38V expression. Scale bars, 50 µm. *p < 0.05, **p < 0.001, ±SEM
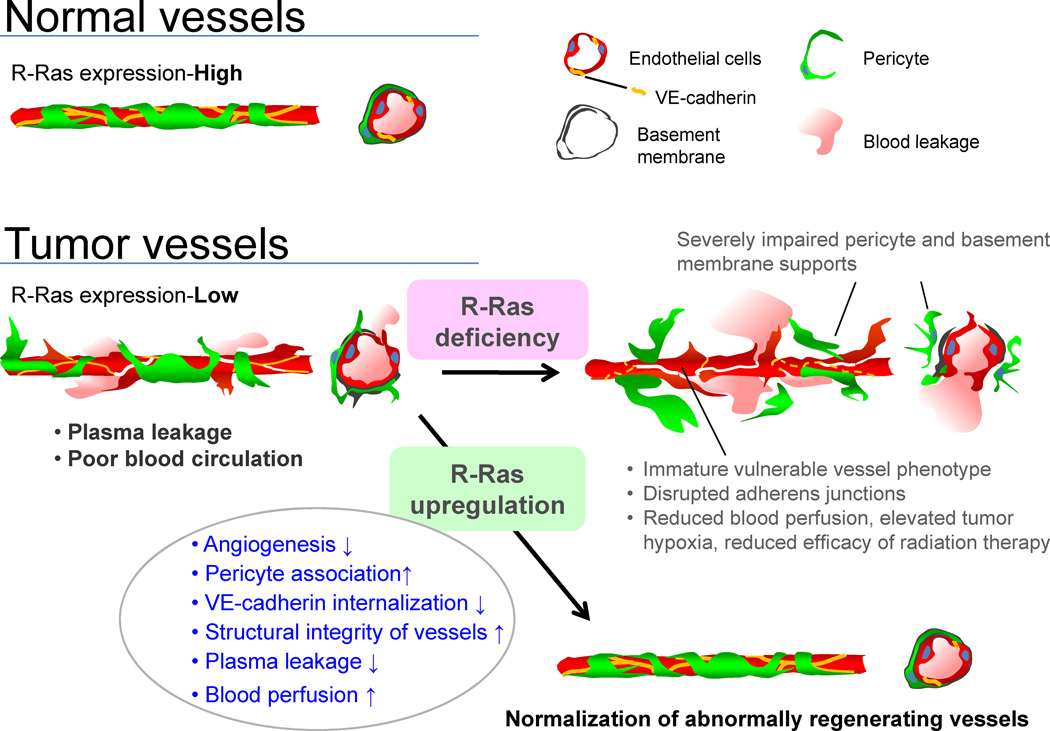
Schematic summary of the consequences of R-Ras disruption and upregulation in tumor blood vessels. The disruption of R-Ras severely impairs structural and functional maturation of tumor vessels. These vessels exhibit the poorly pericyte-supported ‘vulnerable’ immature phenotype, which highly expresses VEGFR2 and αv integrins. The structural abnormalities are associated with extensive blood leakage, reduced blood perfusion, and elevated hypoxia within tumors. On the other hand, upregulation of R-Ras signaling enhances pericyte association and stabilizes VE-cadherin at the endothelial adherens junctions, leading to improved vessel structure and endothelial barrier function with improved blood perfusion. Thus, R-Ras signaling promotes normalization of pathologically regenerating blood vessels.
Comment in
-
Angiogenesis: The new normal.Nat Rev Cancer. 2012 Oct;12(10):660-1. doi: 10.1038/nrc3360. Epub 2012 Sep 6. Nat Rev Cancer. 2012. PMID: 22952010 No abstract available.
References
-
- Allinen M, Beroukhim R, Cai L, Brennan C, Lahti-Domenici J, Huang H, Porter D, Hu M, Chin L, Richardson A, et al. Molecular characterization of the tumor microenvironment in breast cancer. Cancer Cell. 2004;6:17–32. - PubMed
-
- Allingham MJ, van Buul JD, Burridge K. ICAM-1-mediated, Src- and Pyk2-dependent vascular endothelial cadherin tyrosine phosphorylation is required for leukocyte transendothelial migration. J Immunol. 2007;179:4053–4064. - PubMed
-
- Armulik A, Abramsson A, Betsholtz C. Endothelial/pericyte interactions. Circ Res. 2005;97:512–523. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
