

doi: 10.1016/j.devcel.2012.07.011.
Structural architecture and functional evolution of Wnts
Affiliations
- PMID: 22898770
- PMCID: PMC3458506
- DOI: 10.1016/j.devcel.2012.07.011
Item in Clipboard
Structural architecture and functional evolution of Wnts
Dev Cell.
.
Abstract
The crystal structure of a Wnt morphogen bound to its Frizzled receptor ectodomain provides insights into the evolutionary provenance of this complex fold and offers an explanation for why Wnts utilize both lipid- and protein-mediated contacts to engage Frizzleds.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
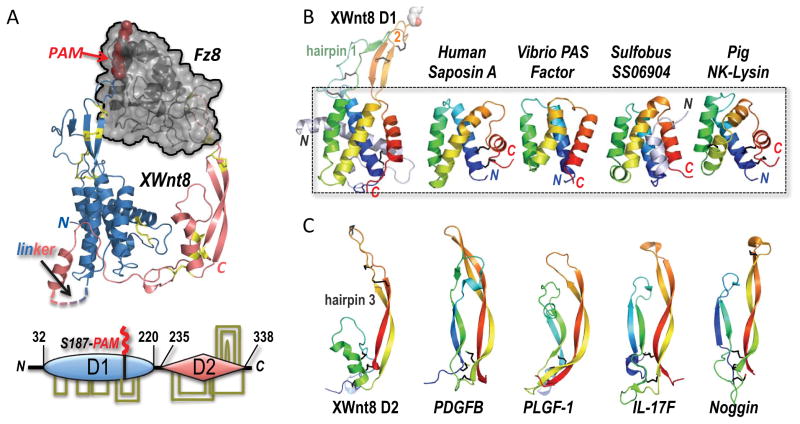
(A) The structure of the binary complex between Xwnt8 and the Fz8 CRD (PDB ID 4F0A) shows that the Wnt utilizes an unusual pair of extended β-hairpin fingers to grasp opposite sides of the (black) Fz domain. The blue N-terminal, mostly helical Wnt domain is lipid-modified (red PAM moiety) at Ser187 at the tip of β-hairpin 2; a hydrophobic groove in the Fz domain sequesters the PAM moiety. The red, mostly β-stranded C-terminal Wnt domain utilizes the tip of another β-hairpin finger to dock a distinct pocket in Fz8. Eleven conserved disulfide bridges are shown in stick format and colored yellow; these links are also drawn in the bottom schematic of the two-domain (D1 and D2) Wnt structure, and fully detailed in Figure 2A. (B) A saposin-like four-α-helix bundle fold forms the core of the D1 domain, and superposes in the 2.85 to 3.9 Å RMSD range to the structures of human Saposin A (PDB ID 2DOB) and Pig NK-Lysin (1NKL)—both conventional saposins with an identifying trio of disulfide bridges, and Vibrio vulnificus PAS factor (PDB ID 2B8I) and Sulfobus solfataricus SS06904 protein (2KJG), examples of atypical, disulfide-free saposins. Hairpins 1 and 2 represent insertions into the first and third interhelical loops, respectively, in the XWnt8 D1 core domain. (C) The structure of the Wnt D2 domain reveals a distinct resemblance to the monomer folds of cystine-knot growth factors, here represented by PDGFB (PDB ID 3MJG), PLGF-1 (PDB ID 1FZV), IL-17F (PDB ID 1JPY) and Noggin (PDB ID 1M4U) (Shim et al., 2010; Iyer et al., 2001; Hymowitz et al., 2001; Groppe et al., 2002). These structures superpose with the XWnt8 D2 domain in the 3 to 3.7 Å RMSD range over 41–46 Cα positions (Krissinel and Henrick, 2004).
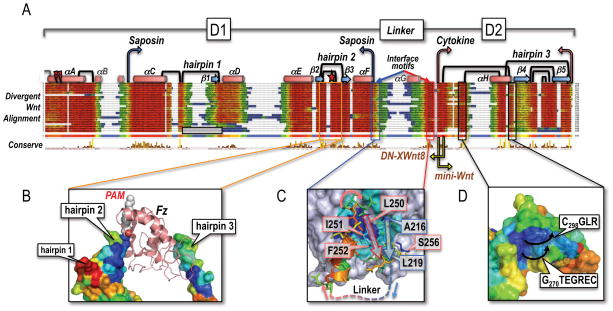
(A) Structure-guided multiple sequence alignment of divergent Wnts to the template structure of XWnt8, and the human Wnt3a chain (Janda et al., 2012). Rather than a uniform set of vertebrate Wnts, we collected very divergent, primitive Wnts from the opposite end of the metazoan spectrum (Pang et al., 2010; Riddiford and Olson, 2011) that would better illustrate the plasticity of the Wnt fold. Sequences were harvested by iterative PsiBLAST searches (Altschul et al., 1997) from Genbank, using as bait the unclassified Wnt subtype chains of available Planaria (Schmidtea mediterranea), Tunicate (Oikopleura dioica), Cnidaria (Nematostella vectensis), Placozoa (Trichoplax adherens), Porifera (Oscarella lobularis and Amphimedon queenslandica) and Ctenophore (Mnemiopsis leidy) genomes; see Supplemental Data for Methods and References. Also included in the screen were the outlier C. elegans Wnts lin-44 and mom-2, and D. melanogaster WntD (Ching et al., 2008). Alignments were captured in Jalview with the ClustalX color scheme (Waterhouse et al., 2009); the consensus sequence logo is displayed in pink. Helices as drawn as red cylinders and labeled αA-αH (similar to the nomenclature of Janda et al., 2012), while β-strands are depicted as blue arrows and called β1-β5 (different from Janda et al., 2012). The boundaries of the saposin-like D1 and cytokine-like D2 domains are noted with blue and pink arrows, respectively; yellow arrows mark the N-terminal end of the Janda et al. (2012) mini-Wnt construct, and the C-terminal boundary of the DN-XWnt8 truncation (Hoppler et al., 1996). The Tiki1 processing sites in the XWnt8 helix αA are indicated by red lightning bolts (Zhang et al., 2012). The eleven X-ray-defined disulfide bridges (Figure 1A) are drawn as black links; an N-terminal grey half-link notes a likely, additional disulfide bridge in some Wnts, but absent in XWnt8. The three hairpin structures are also numbered, with the lipidated Ser187 (marked with a red star) located in hairpin 2, and the vicinal disulfide bridge in hairpin 3. Hairpin 1 is notably absent in the Oikopleura Wnts, as noted by a grey, empty box. The length and sequence-variable linker region between D1 and D2 domains is bordered by well conserved interface motifs that closely associate in a near parallel β-strand fashion at the base of the saposin-like helical bundle; the intervening sequence is largely unstructured (and not visible in the XWnt8 structure, save for a short helix αG). (B) The multiple alignment of deeply divergent Wnts was used to color the molecular surface of the XWnt8 structure using the ConSurf program (Ashkenazy et al., 2010), showing nearly invariant (blue) to completely unconserved (red) sequence patches. The present view is of the the captured Fz domain, with a highly divergent (and collapsed) hairpin 1, a very well conserved hairpin 2 (lipidated with the PAM group bound by Fz; orange boxed region), and only a moderately conserved hairpin 3, that is involved in protein-protein contacts with the second Fz binding site. (C) Close-up view of the two conserved interface motifs in blue (D1 domain) and red (D2 domain) that associate in a parallel fashion; blue and red arrows illustrate the path of the underlying chain. The missing linker region is noted by a dotted line. Key conserved residues in XWnt8 are boxed and numbered. (D) A conserved, convex epitope at the base of the D2 domain in Wnts that may indicate the interaction site with LRP5/6, by analogy with the sclerostin contact site (Bourhis et al., 2011; Bao et al., 2012). The black arrows indicate the direction of the two loops in XWnt8 that respectively follow Gly270 and Cys298.
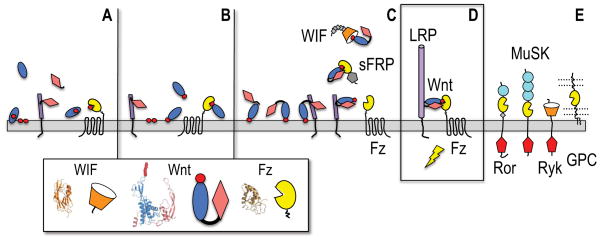
(A) A membrane-interacting saposin-like domain (blue oval) has a (red circle) lipid-loading role for a Fz-like GPCR. A secreted cystine knot cytokine (pink diamond) binds a cell surface LRP-like progenitor (purple cylinder). (B) The saposin-like domain gains a Ser-acylation site in its membrane-interacting loops and can then directly bind the Fz domain of the GPCR. (C) A hydrophobic motif in the cytokine N-terminal extension can dock to the saposin helical domain (Figure S1), and a fortuitous gene fusion of these interacting proteins creates a two domain Wnt progenitor that retains cytokine-mediated binding to a primitive LRP, and can be presented together to the Fz GPCR principally by lipid docking. They cytokine portion of the Wnt has an evolved affinity for the interacting Fz and WIF domains, on the receptors or soluble binding proteins (sFRPs, WIF). Small grey diamonds mark the EGF repeat tail in WIF, while a grey pentagon in sFRP illustrates a C-terminal netrin module. (D) The Wnt remains the critical bridging element in the active ternary receptor complex with LRP5/6 and a Fz GPCR. The purple cylinder in the LRP chain (or its shorter progenitor LRP) denotes the four-fold-repeat of β-propeller and EGF domains. (E) Additional signaling receptors (Ror, MuSK and Ryk) and presentation molecules (glypican or GPC) can potentially engage lipidated Wnts. Light blue circles in Ror and MuSK ectodomains mark Ig modules, while intracellular tyrosine kinases are noted by red pentagons.
References
-
- Bazan JF, de Sauvage FJ. Cell. 2009;138:1055–1056. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
