

Post-translational modification: nature's escape from genetic imprisonment and the basis for dynamic information encoding
- PMID: 22899623
- PMCID: PMC3473174
- DOI: 10.1002/wsbm.1185
Post-translational modification: nature's escape from genetic imprisonment and the basis for dynamic information encoding
Abstract
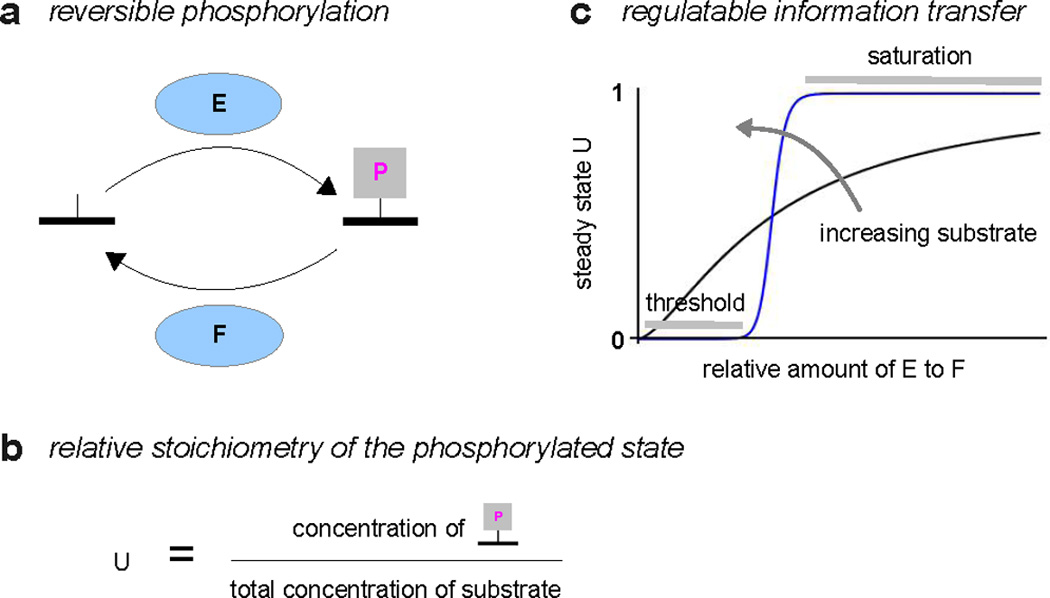
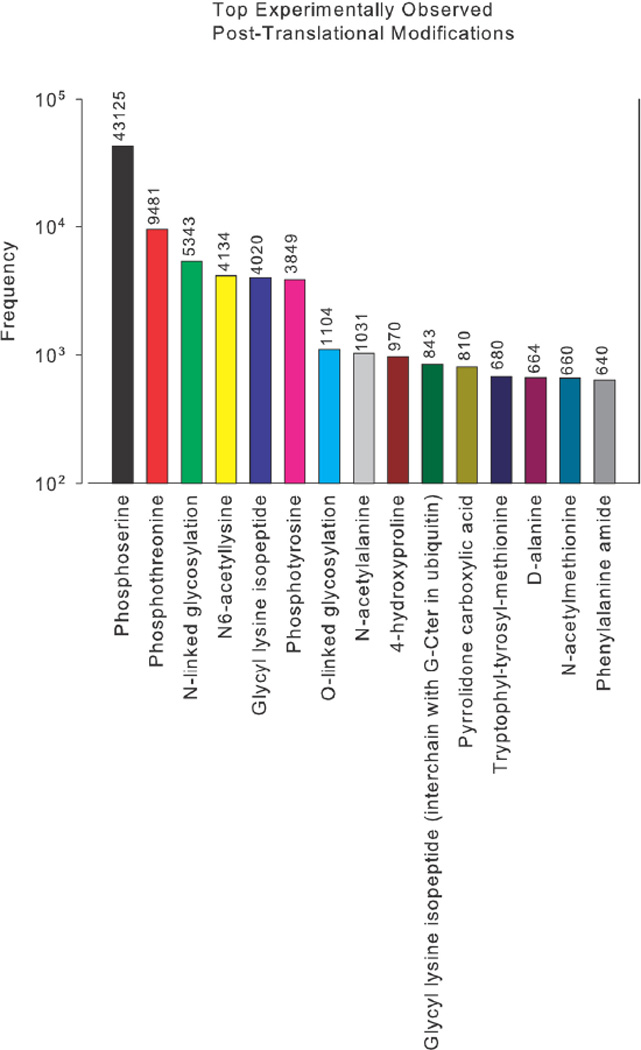
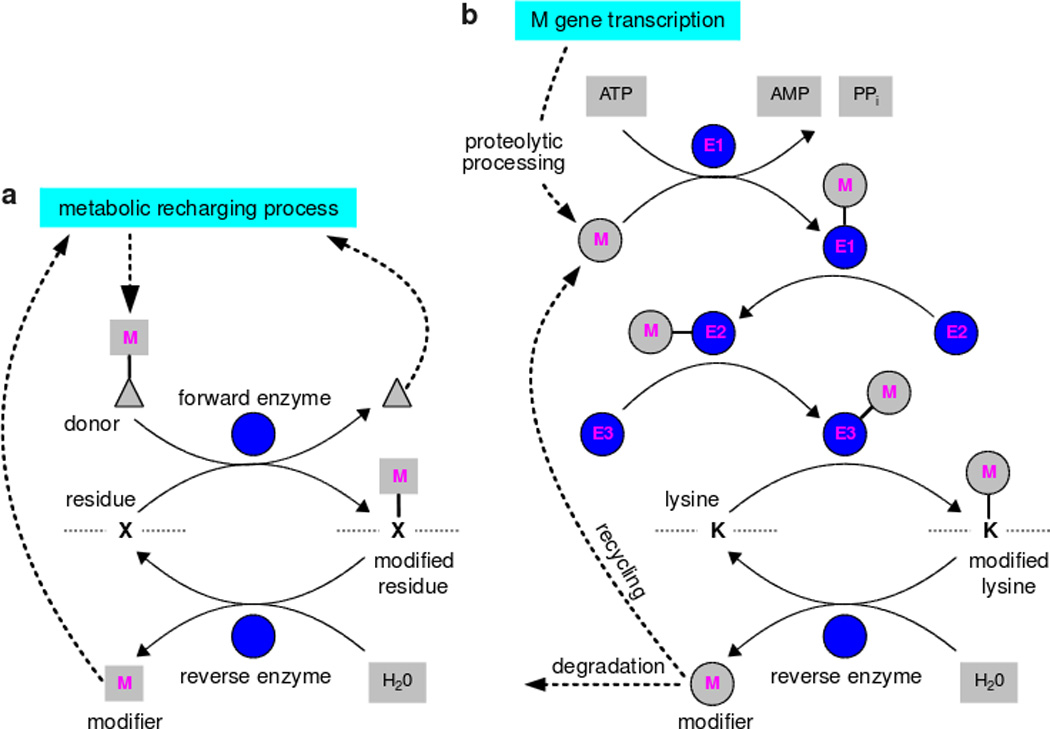
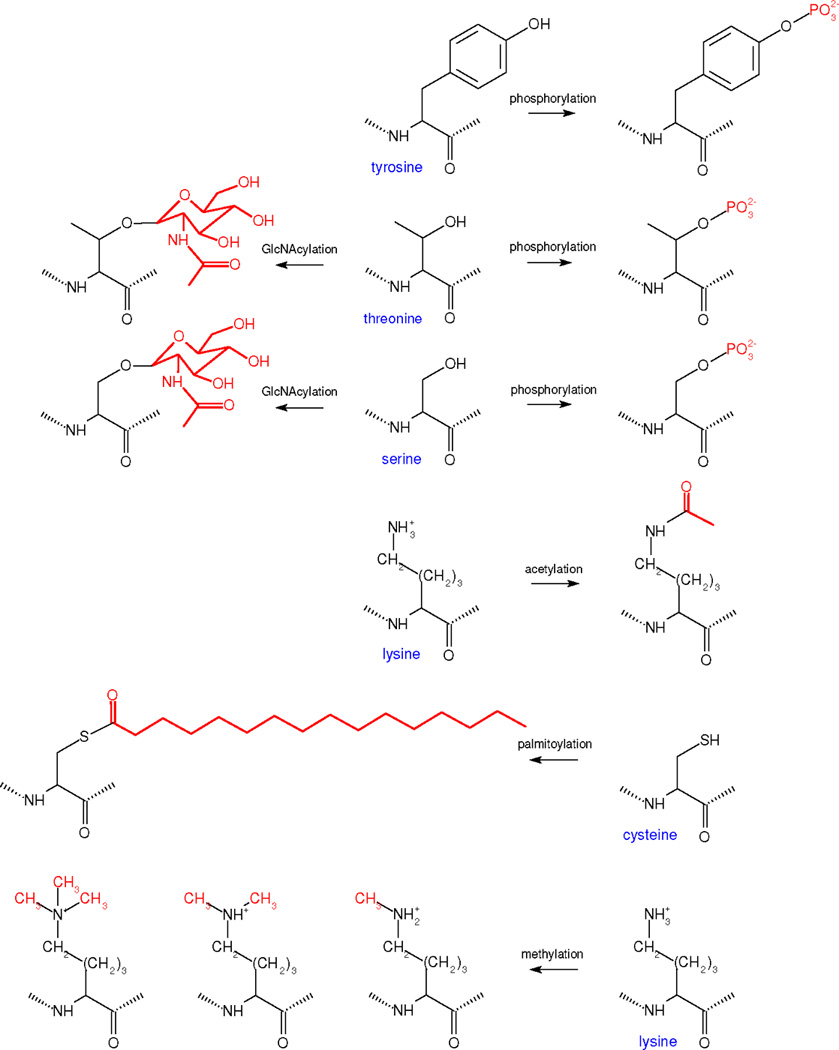
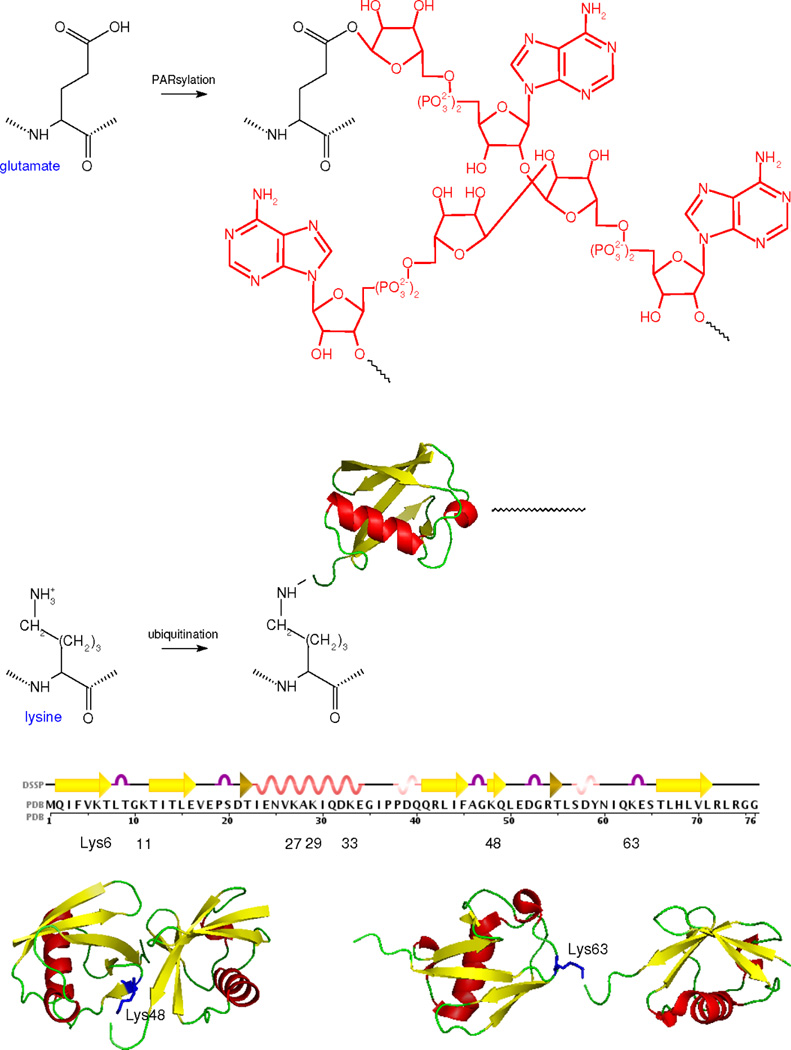
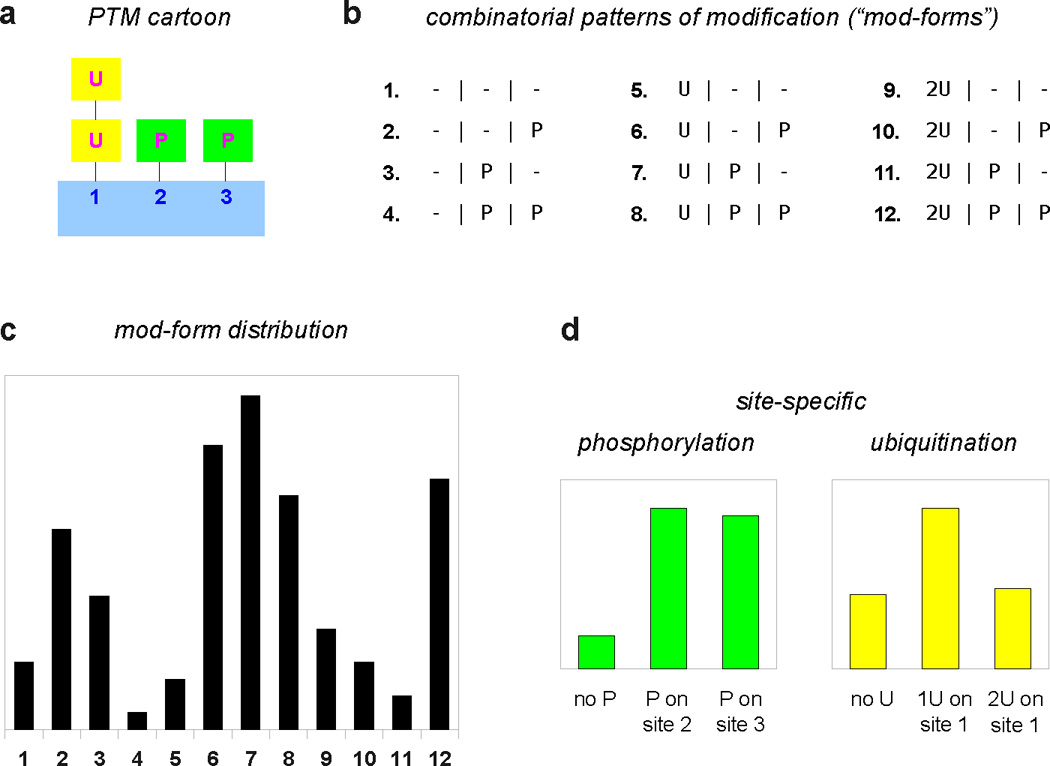
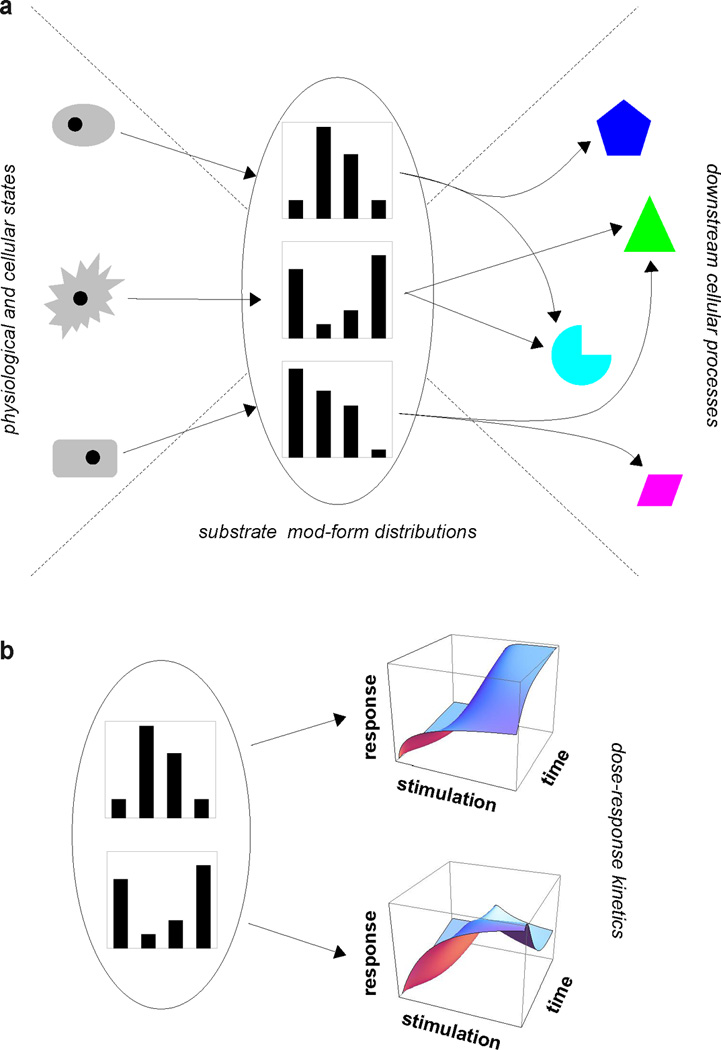
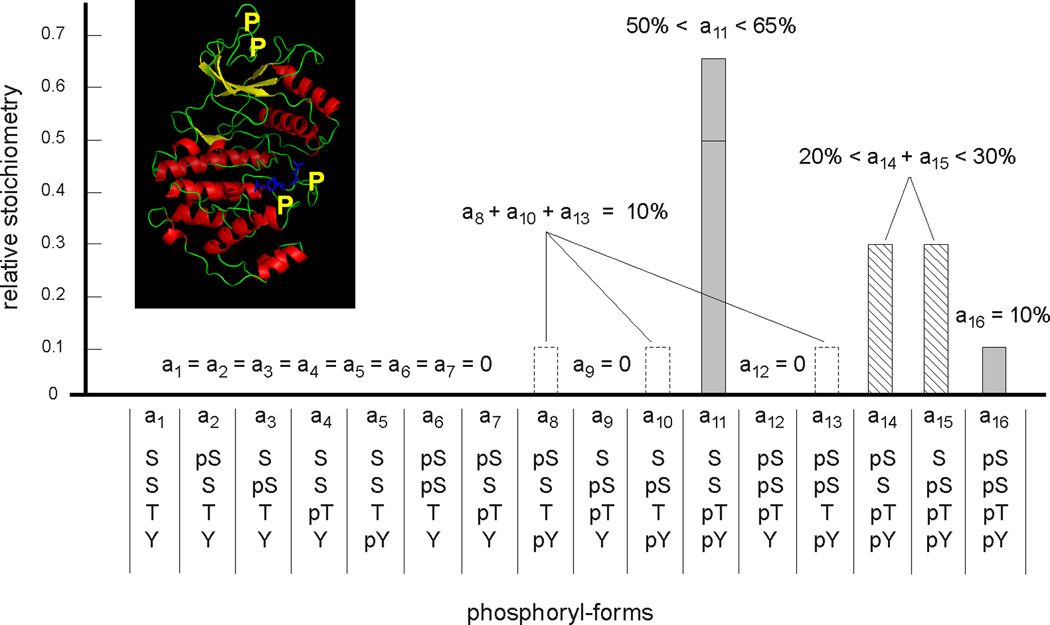
We discuss protein post-translational modification (PTM) from an information processing perspective. PTM at multiple sites on a protein creates a combinatorial explosion in the number of potential 'mod-forms', or global patterns of modification. Distinct mod-forms can elicit distinct downstream responses, so that the overall response depends partly on the effectiveness of a particular mod-form to elicit a response and partly on the stoichiometry of that mod-form in the molecular population. We introduce the 'mod-form distribution'-the relative stoichiometries of each mod-form-as the most informative measure of a protein's state. Distinct mod-form distributions may summarize information about distinct cellular and physiological conditions and allow downstream processes to interpret this information accordingly. Such information 'encoding' by PTMs may facilitate evolution by weakening the need to directly link upstream conditions to downstream responses. Mod-form distributions provide a quantitative framework in which to interpret ideas of 'PTM codes' that are emerging in several areas of biology, as we show by reviewing examples of ion channels, GPCRs, microtubules, and transcriptional co-regulators. We focus particularly on examples other than the well-known 'histone code', to emphasize the pervasive use of information encoding in molecular biology. Finally, we touch briefly on new methods for measuring mod-form distributions.
Copyright © 2012 Wiley Periodicals, Inc.
Figures
References
-
- Acuto O, Di Bartolo v, Michel F. Tailoring T-cell receptor signals by proximal negative feedback mechanisms. Nat. Rev. Immunol. 2008;8:699–712. - PubMed
-
- Al-Hakim AK, Zagorska A, Chapman L, Deak M, Peggie M, Alessi DR. Control of AMPK-related kinases by USP9X and atypical Lys29/Lys33-linked polyubiquitin chains. Biochem. J. 2008;411:249–260. - PubMed
-
- Alvarez-Gonzalez R, Jacobson MK. Characterization of polymers of adenosine diphosphate ribose generated in vitro and in vivo. Biochemistry. 1987;26:3218–3224. - PubMed
-
- Amerik AY, Hochstrasser M. Mechanism and function of deubiquitinating enzymes. Biochim. Biophys. Acta. 2004;1695:189–207. - PubMed
-
- Amniai L, Barbier P, Sillen A, Wieruszeski J-M, Peyrot v, Lippens G, Landrieu I. Alzheimer disease specific phosphoepitopes of Tau interfere with assembly of tubulin but not binding to microtubules. FASEB J. 2009;23:1146–1152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
